Tong Lei, Denu John M
Department of Biomolecular Chemistry, University of Wisconsin, School of Medicine and Public Health, Madison, WI 53706, USA.
Biochim Biophys Acta. 2010 Aug;1804(8):1617-25. doi: 10.1016/j.bbapap.2010.02.007. Epub 2010 Feb 20.
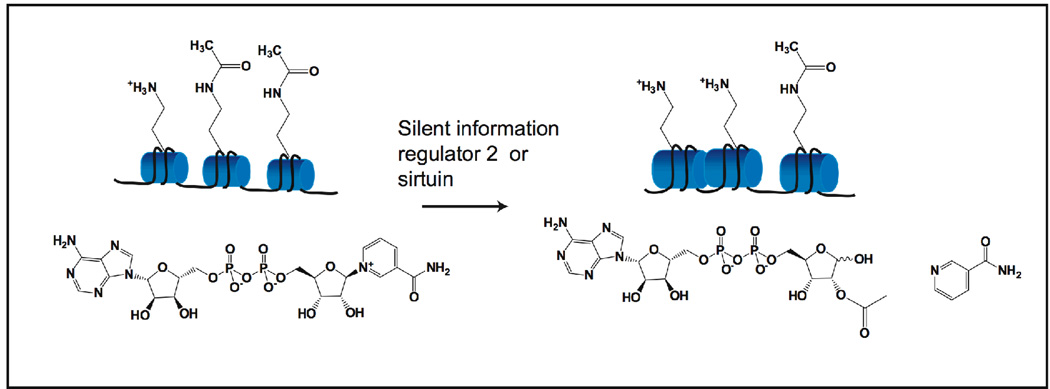
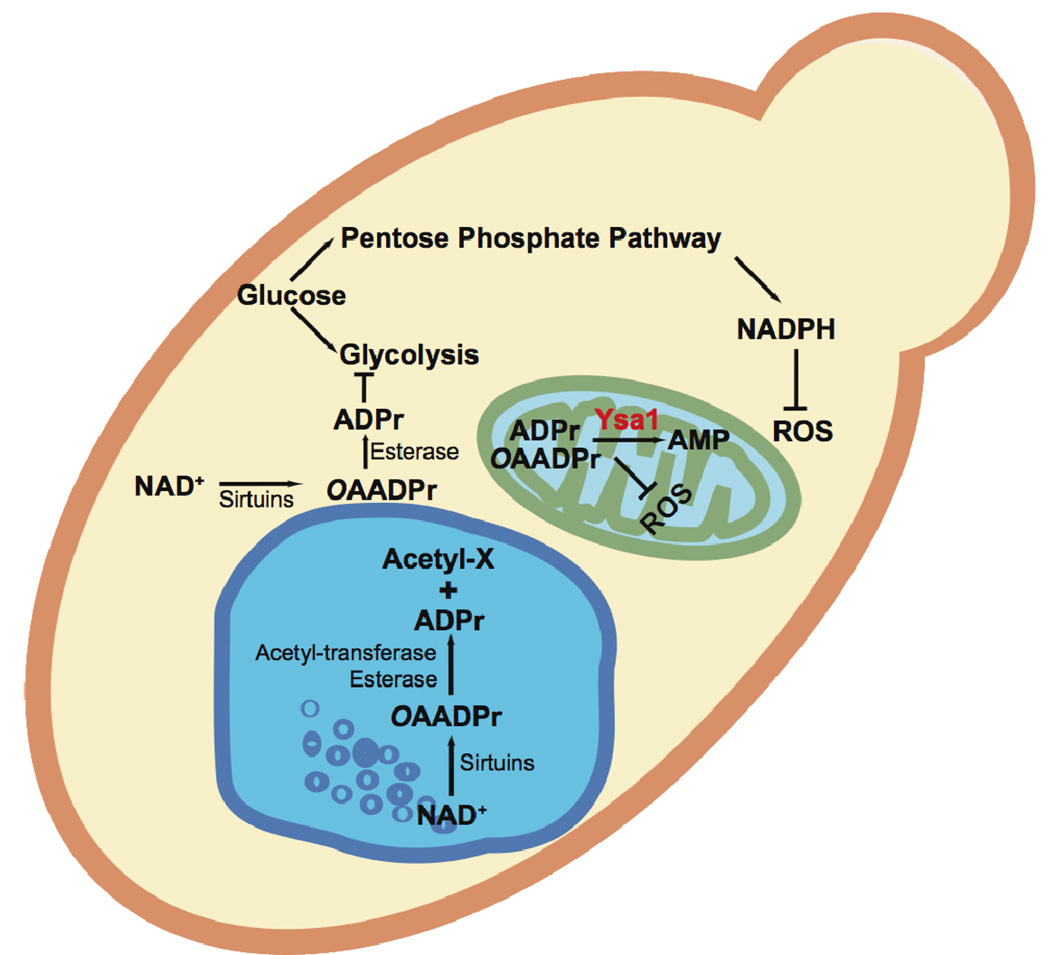
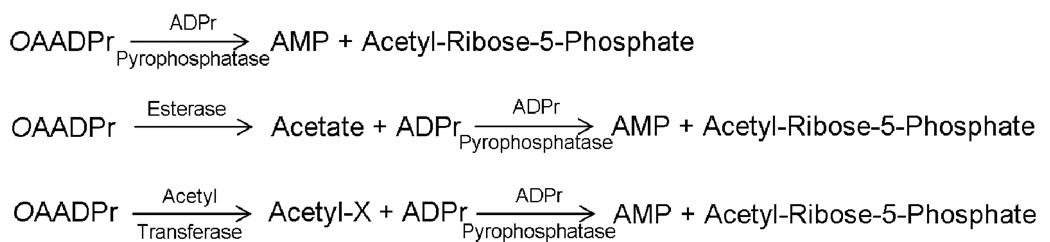
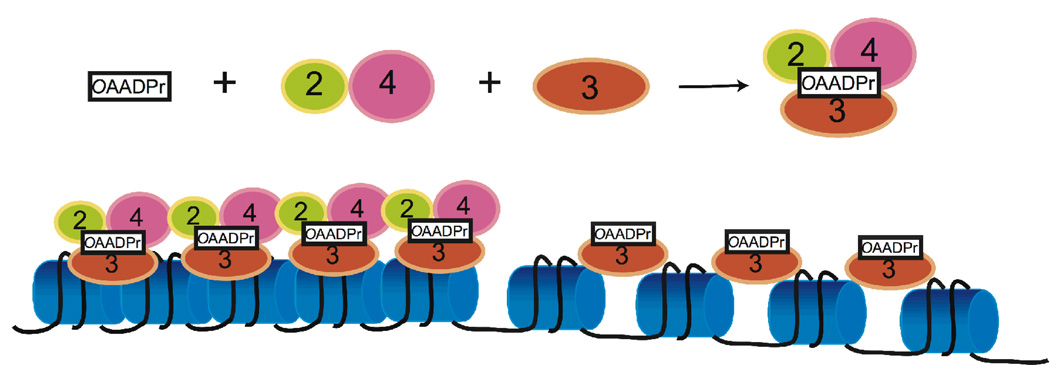
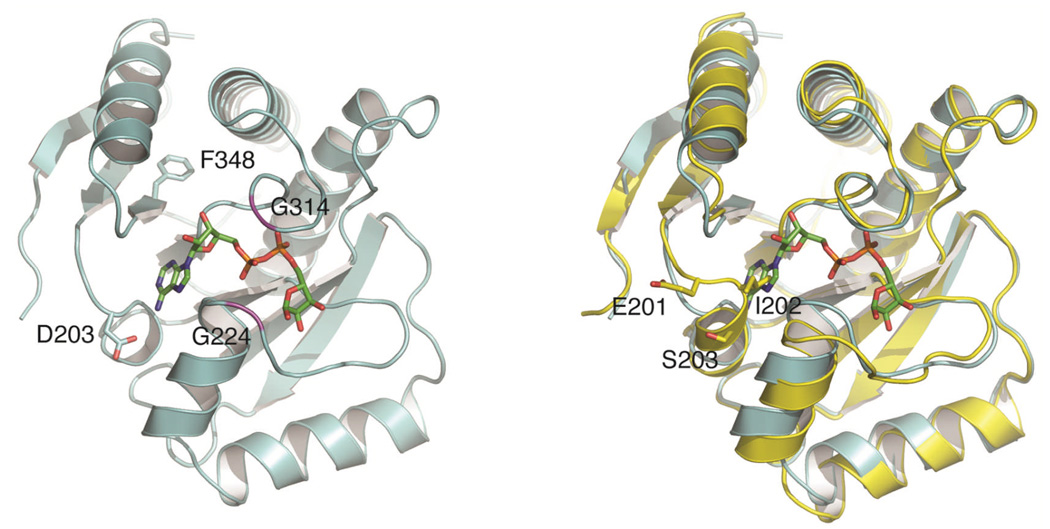
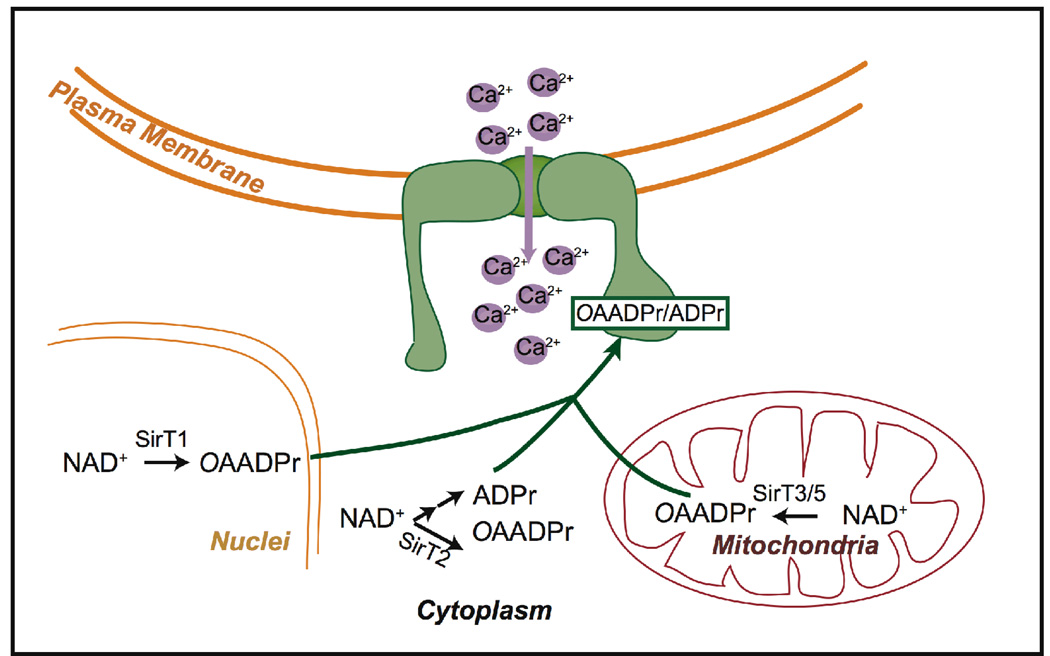
Sirtuins catalyze the NAD(+)-dependent deacetylation of target proteins, which are regulated by this reversible lysine modification. During deacetylation, the glycosidic bond of the nicotinamide ribose is cleaved to yield nicotinamide and the ribose accepts the acetyl group from substrate to produce O-acetyl-ADP-ribose (OAADPr), which exists as an approximately 50:50 mixture of 2' and 3' isomers at neutral pH. Discovery of this metabolite has fueled the idea that OAADPr may play an important role in the biology associated with sirtuins, acting as a signaling molecule and/or an important substrate for downstream enzymatic processes. Evidence for OAADPr-metabolizing enzymes indicates that at least three distinct activities exist that could modulate the cellular levels of this NAD(+)-derived metabolite. In Saccharomyces cerevisiae, NUDIX hydrolase Ysa1 cleaves OAADPr to AMP and 2- and 3-O-acetylribose-5-phosphate, lowering the cellular levels of OAADPr. A buildup of OAADPr and ADPr has been linked to a metabolic shift that lowers endogenous reactive oxygen species and diverts glucose towards preventing oxidative damage. In vitro, the mammalian enzyme ARH3 hydrolyzes OAADPr to acetate and ADPr. A third nuclear-localized activity appears to utilize OAADPr to transfer the acetyl-group to another small molecule, whose identity remains unknown. Recent studies suggest that OAADPr may regulate gene silencing by facilitating the assembly and loading of the Sir2-4 silencing complex onto nucleosomes. In mammalian cells, the Trpm2 cation channel is gated by both OAADPr and ADP-ribose. Binding is mediated by the NUDIX homology (NudT9H) domain found within the intracellular portion of the channel. OAADPr is capable of binding the Macro domain of splice variants from histone protein MacroH2A, which is highly enriched at heterochromatic regions. With recently developed tools, the pace of new discoveries of OAADPr-dependent processes should facilitate new molecular insight into the diverse biological processes modulated by sirtuins.
沉默调节蛋白催化靶蛋白的NAD⁺依赖性去乙酰化,而靶蛋白受这种可逆的赖氨酸修饰调控。在去乙酰化过程中,烟酰胺核糖的糖苷键断裂生成烟酰胺,核糖从底物接受乙酰基生成O-乙酰-ADP-核糖(OAADPr),在中性pH条件下,OAADPr以2'和3'异构体约50:50的混合物形式存在。这种代谢物的发现引发了一种观点,即OAADPr可能在与沉默调节蛋白相关的生物学过程中发挥重要作用,充当信号分子和/或下游酶促过程的重要底物。OAADPr代谢酶的证据表明,至少存在三种不同的活性,可调节这种NAD⁺衍生代谢物的细胞水平。在酿酒酵母中,NUDIX水解酶Ysa1将OAADPr裂解为AMP以及2-和3-O-乙酰核糖-5-磷酸,从而降低细胞内OAADPr的水平。OAADPr和ADPr的积累与一种代谢转变有关,这种转变会降低内源性活性氧水平,并将葡萄糖转向预防氧化损伤。在体外,哺乳动物酶ARH3将OAADPr水解为乙酸盐和ADPr。第三种定位于细胞核的活性似乎利用OAADPr将乙酰基转移到另一个小分子上,该小分子的身份仍然未知。最近的研究表明,OAADPr可能通过促进Sir2-4沉默复合物组装并加载到核小体上来调节基因沉默。在哺乳动物细胞中,瞬时受体电位阳离子通道蛋白2(Trpm2)阳离子通道由OAADPr和ADP-核糖门控。结合由通道细胞内部分的NUDIX同源(NudT9H)结构域介导。OAADPr能够结合组蛋白MacroH2A剪接变体的Macro结构域,该结构域在异染色质区域高度富集。借助最近开发的工具,依赖OAADPr的过程的新发现速度应有助于对沉默调节蛋白调节的多种生物学过程有新的分子见解。