Viral Infections Unit, Department of Vaccination and Immune Protection, National Institute for Health and Welfare THL, Helsinki, Finland.
PLoS One. 2010 Oct 20;5(10):e13329. doi: 10.1371/journal.pone.0013329.
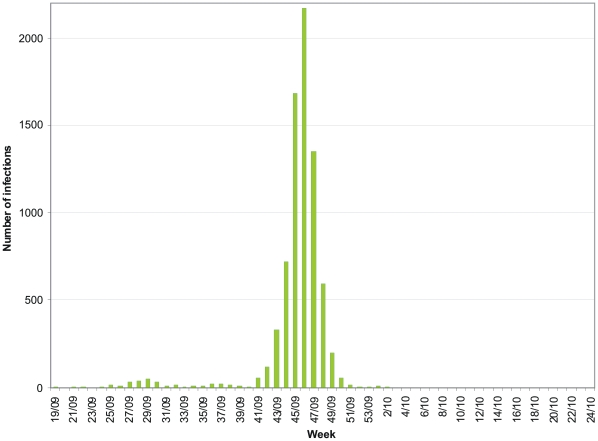
In Finland, the first infections caused by the 2009 pandemic influenza A(H1N1) virus were identified on May 10. During the next three months almost all infections were found from patients who had recently traveled abroad. In September 2009 the pandemic virus started to spread in the general population, leading to localized outbreaks and peak epidemic activity was reached during weeks 43-48.
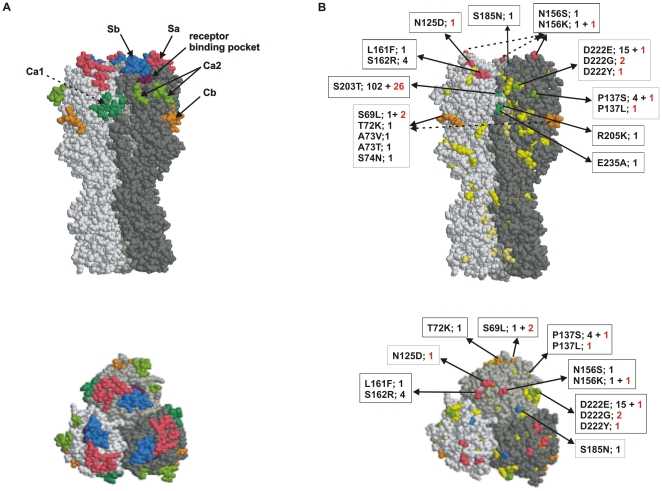
METHODS/RESULTS: The nucleotide sequences of the hemagglutinin (HA) and neuraminidase (NA) genes from viruses collected from 138 patients were determined. The analyzed viruses represented mild and severe infections and different geographic regions and time periods. Based on HA and NA gene sequences, the Finnish pandemic viruses clustered in four groups. Finnish epidemic viruses and A/California/07/2009 vaccine virus strain varied from 2-8 and 0-5 amino acids in HA and NA molecules, respectively, giving a respective maximal evolution speed of 1.4% and 1.1%. Most amino acid changes in HA and NA molecules accumulated on the surface of the molecule and were partly located in antigenic sites. Three severe infections were detected with a mutation at HA residue 222, in two viruses with a change D222G, and in one virus D222Y. Also viruses with change D222E were identified. All Finnish pandemic viruses were sensitive to oseltamivir having the amino acid histidine at residue 275 of the neuraminidase molecule.
The Finnish pandemic viruses were quite closely related to A/California/07/2009 vaccine virus. Neither in the HA nor in the NA were changes identified that may lead to the selection of a virus with increased epidemic potential or exceptionally high virulence. Continued laboratory-based surveillance of the 2009 pandemic influenza A(H1N1) is important in order to rapidly identify drug resistant viruses and/or virus variants with potential ability to cause severe forms of infection and an ability to circumvent vaccine-induced immunity.
2009 年甲型 H1N1 流感大流行病毒在芬兰的首次感染病例于 5 月 10 日确诊。在接下来的三个月中,几乎所有的感染病例都来自近期有出国旅行史的患者。2009 年 9 月,大流行病毒开始在普通人群中传播,导致局部暴发,第 43-48 周达到疫情高峰。
方法/结果:对从 138 名患者中采集的病毒的血凝素(HA)和神经氨酸酶(NA)基因的核苷酸序列进行了测定。分析的病毒代表了轻症和重症感染,以及不同的地理区域和时间。根据 HA 和 NA 基因序列,芬兰大流行病毒分为 4 组。芬兰流行病毒株和 A/California/07/2009 疫苗病毒株在 HA 和 NA 分子上分别有 2-8 个和 0-5 个氨基酸差异,相应的最大进化速度分别为 1.4%和 1.1%。HA 和 NA 分子表面的大多数氨基酸变化部分位于抗原位点。在 3 例重症感染中检测到 HA 残基 222 处的突变,在 2 株病毒中为 D222G,在 1 株病毒中为 D222Y。也发现了 D222E 改变的病毒。所有芬兰大流行病毒均对奥司他韦敏感,神经氨酸酶分子上的残基 275 具有组氨酸。
芬兰大流行病毒与 A/California/07/2009 疫苗病毒非常密切相关。在 HA 和 NA 中均未发现可能导致流行潜力增加或异常高毒力的病毒的变化。为了快速识别具有耐药性的病毒和/或可能导致严重感染形式和规避疫苗诱导免疫能力的病毒变异,继续进行基于实验室的 2009 年甲型 H1N1 流感大流行的监测非常重要。