Tamamaki Nobuaki, Tomioka Ryohei
Department of Morphological Neural Science, Graduate School of Medical Sciences, Kumamoto University Kumamoto, Japan.
Front Neurosci. 2010 Dec 8;4:202. doi: 10.3389/fnins.2010.00202. eCollection 2010.
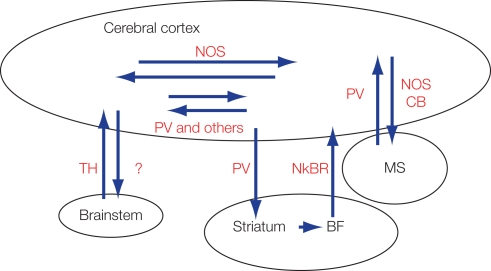
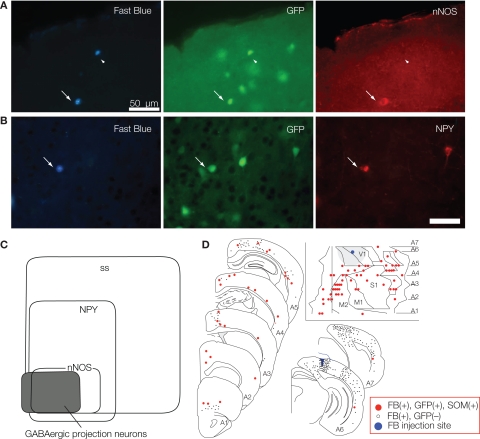
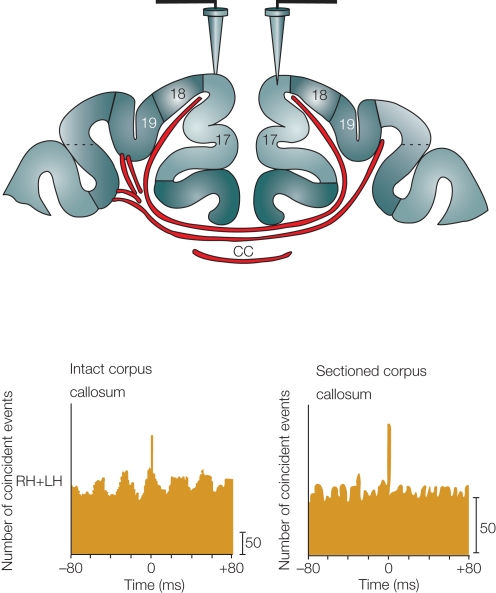
Features and functions of long-range GABAergic projection neurons in the developing cerebral cortex have been reported previously, although until now their significance in the adult cerebral cortex has remained uncertain. The septo-hippocampal circuit is one exception - in this system, long-range mature GABAergic projection neurons have been well analyzed and their contribution to the generation of theta-oscillatory behavior in the hippocampus has been documented. To have a clue to the function of the GABAergic projection neurons in the neocortex, we view how the long-range GABAergic projections are integrated in the cortico-cortical, cortico-fugal, and afferent projections in the cerebral cortex. Then, we consider the possibility that the GABAergic projection neurons are involved in the generation, modification, and/or synchronization of oscillations in mature neocortical neuron activity. When markers that identify the GABAergic projection neurons are examined in anatomical and developmental studies, it is clear that neuronal NO synthetase (nNOS)-immunoreactivity can readily identify GABAergic projection neurons. GABAergic projection neurons account for 0.5% of the neocortical GABAergic neurons. To elucidate the role of the GABAergic projection neurons in the neocortex, it will be necessary to clarify the network constructed by nNOS-positive GABAergic projection neurons and their postsynaptic targets. Thus, our long-range goals will be to label and manipulate (including deleting) the GABAergic projection neurons using genetic tools driven by a nNOS promoter. We recognize that this may be a complex endeavor, as most excitatory neurons in the murine neocortex express nNOS transiently. Nevertheless, additional studies characterizing long-range GABAergic projection neurons will have great value to the overall understanding of mature cortical function.
发育中的大脑皮质中长距离GABA能投射神经元的特征和功能此前已有报道,尽管迄今为止它们在成人大脑皮质中的意义仍不明确。隔区-海马回路是一个例外——在这个系统中,长距离成熟GABA能投射神经元已得到充分分析,并且它们对海马体中θ振荡行为产生的贡献也已被记录。为了了解新皮质中GABA能投射神经元的功能,我们观察了长距离GABA能投射是如何整合到大脑皮质的皮质-皮质、皮质-传出和传入投射中的。然后,我们考虑了GABA能投射神经元参与成熟新皮质神经元活动振荡的产生、调节和/或同步的可能性。当在解剖学和发育研究中检查识别GABA能投射神经元的标记物时,很明显神经元型一氧化氮合酶(nNOS)免疫反应性能够轻易识别GABA能投射神经元。GABA能投射神经元占新皮质GABA能神经元的0.5%。为了阐明GABA能投射神经元在新皮质中的作用,有必要弄清楚由nNOS阳性GABA能投射神经元及其突触后靶点构建的网络。因此,我们的长期目标是利用由nNOS启动子驱动的基因工具标记和操纵(包括删除)GABA能投射神经元。我们认识到这可能是一项复杂的工作,因为小鼠新皮质中的大多数兴奋性神经元会短暂表达nNOS。尽管如此,对长距离GABA能投射神经元进行的更多特征研究将对全面理解成熟皮质功能具有重要价值。