Division of Mathematics, University of Dundee, Dundee, United Kingdom.
PLoS One. 2011 Feb 28;6(2):e16980. doi: 10.1371/journal.pone.0016980.
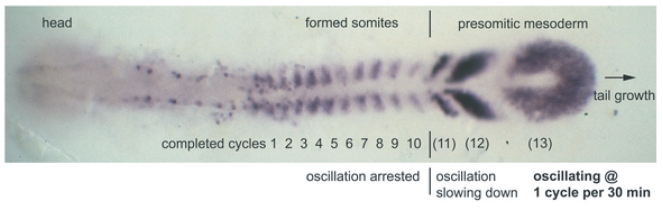
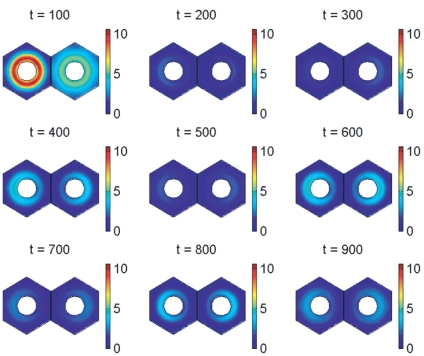
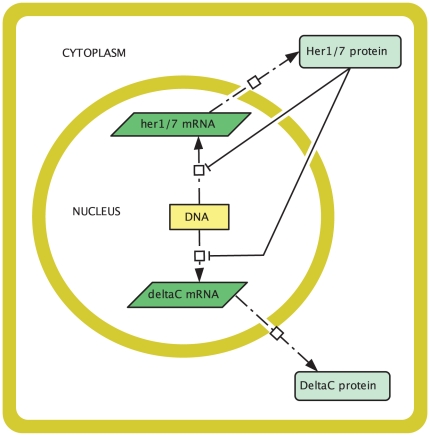
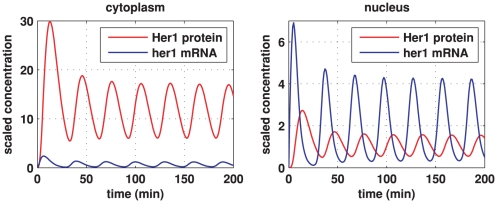
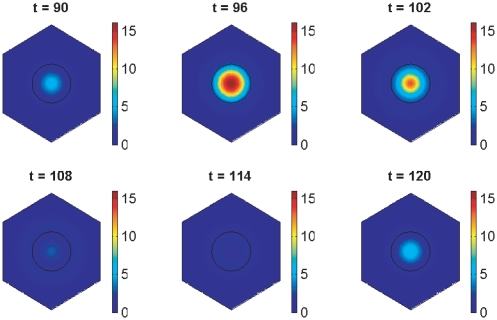
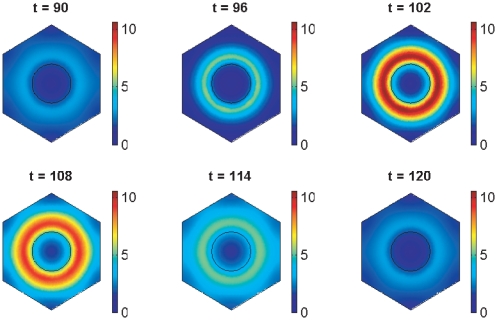
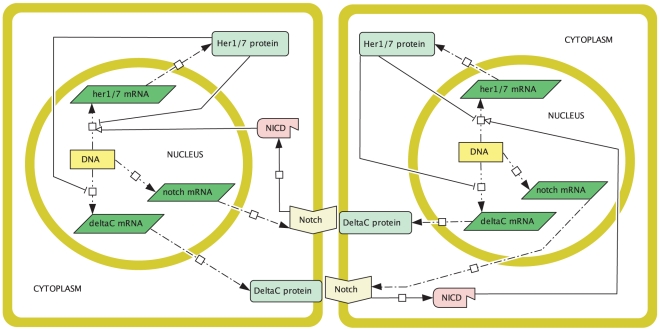
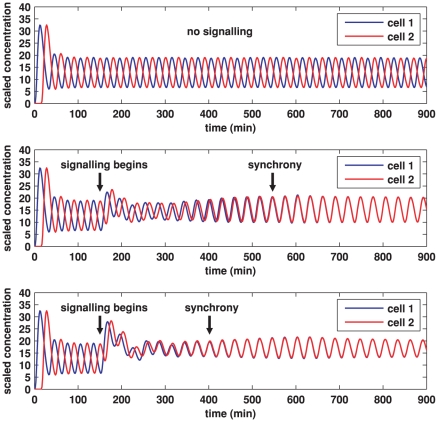
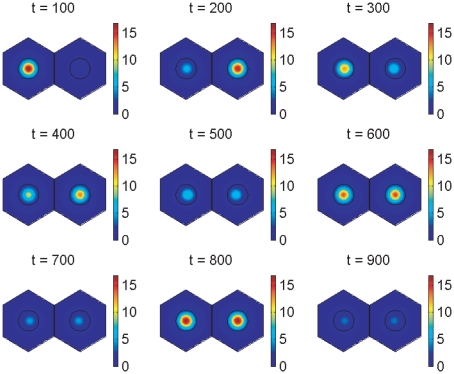
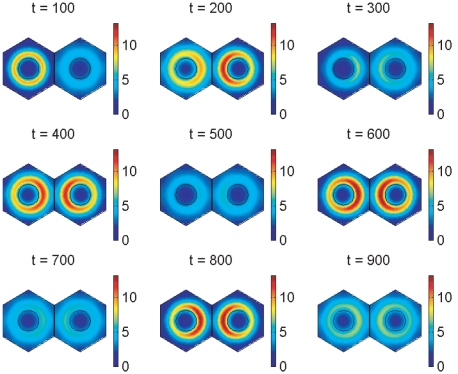
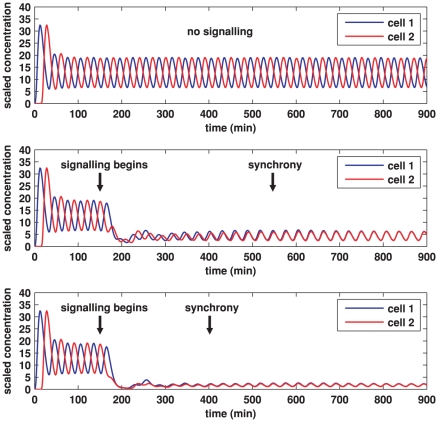
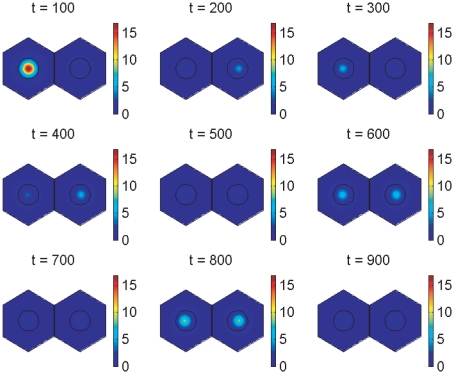
In the vertebrate embryo, tissue blocks called somites are laid down in head-to-tail succession, a process known as somitogenesis. Research into somitogenesis has been both experimental and mathematical. For zebrafish, there is experimental evidence for oscillatory gene expression in cells in the presomitic mesoderm (PSM) as well as evidence that Notch signalling synchronises the oscillations in neighbouring PSM cells. A biological mechanism has previously been proposed to explain these phenomena. Here we have converted this mechanism into a mathematical model of partial differential equations in which the nuclear and cytoplasmic diffusion of protein and mRNA molecules is explicitly considered. By performing simulations, we have found ranges of values for the model parameters (such as diffusion and degradation rates) that yield oscillatory dynamics within PSM cells and that enable Notch signalling to synchronise the oscillations in two touching cells. Our model contains a Hill coefficient that measures the co-operativity between two proteins (Her1, Her7) and three genes (her1, her7, deltaC) which they inhibit. This coefficient appears to be bounded below by the requirement for oscillations in individual cells and bounded above by the requirement for synchronisation. Consistent with experimental data and a previous spatially non-explicit mathematical model, we have found that signalling can increase the average level of Her1 protein. Biological pattern formation would be impossible without a certain robustness to variety in cell shape and size; our results possess such robustness. Our spatially-explicit modelling approach, together with new imaging technologies that can measure intracellular protein diffusion rates, is likely to yield significant new insight into somitogenesis and other biological processes.
在脊椎动物胚胎中,组织块称为体节,以头尾相继的方式形成,这个过程称为体节形成。体节形成的研究既有实验性的,也有数学性的。对于斑马鱼,有实验证据表明,在原条中胚层(PSM)细胞中存在基因表达的振荡,并且 Notch 信号能够使相邻 PSM 细胞的振荡同步。之前已经提出了一种生物学机制来解释这些现象。在这里,我们将这个机制转化为一个偏微分方程的数学模型,其中明确考虑了蛋白质和 mRNA 分子的核和细胞质扩散。通过进行模拟,我们找到了模型参数(如扩散和降解速率)的范围值,这些值在 PSM 细胞中产生了振荡动力学,并使 Notch 信号能够使两个接触的细胞的振荡同步。我们的模型包含一个 Hill 系数,用于衡量两种蛋白质(Her1、Her7)和它们抑制的三个基因(her1、her7、deltaC)之间的合作性。这个系数似乎受到两个方面的限制:一方面是单个细胞中振荡的要求,另一方面是同步的要求。与实验数据和以前的非空间显式数学模型一致,我们发现信号可以增加 Her1 蛋白的平均水平。如果没有对细胞形状和大小的变化具有一定的稳健性,生物模式形成将是不可能的;我们的结果具有这种稳健性。我们的空间显式建模方法,以及可以测量细胞内蛋白质扩散速率的新成像技术,可能会对体节形成和其他生物学过程产生重大的新见解。