Department of Physiology and Biophysics, Dalhousie University, Halifax, Nova Scotia B3H 1X5, Canada.
J Gen Physiol. 2011 Aug;138(2):165-78. doi: 10.1085/jgp.201110605. Epub 2011 Jul 11.
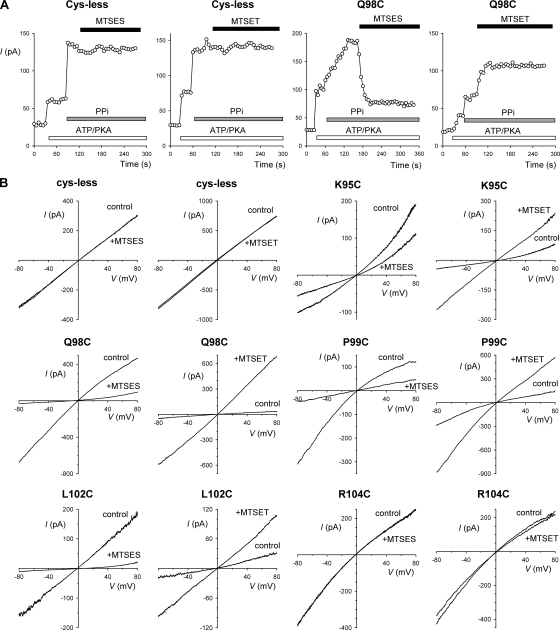
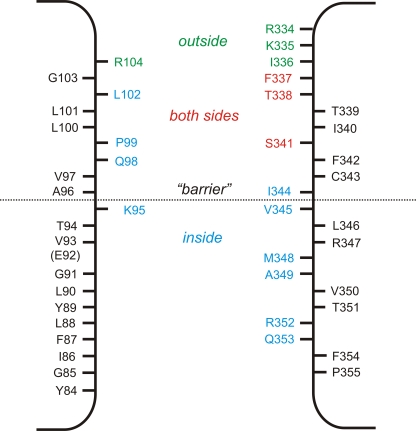
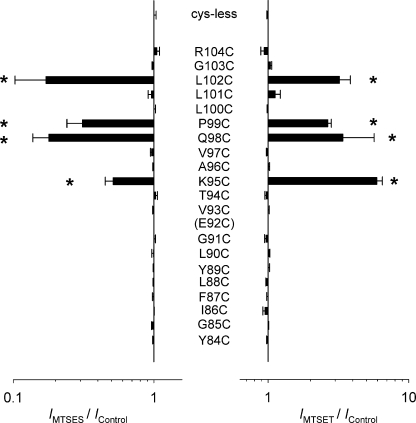
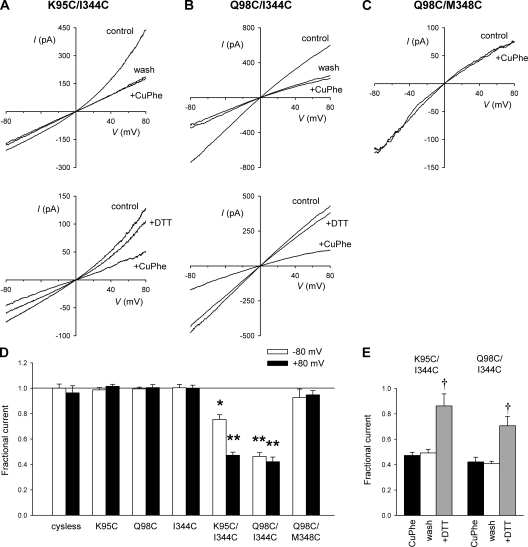
Different transmembrane (TM) α helices are known to line the pore of the cystic fibrosis TM conductance regulator (CFTR) Cl(-) channel. However, the relative alignment of these TMs in the three-dimensional structure of the pore is not known. We have used patch-clamp recording to investigate the accessibility of cytoplasmically applied cysteine-reactive reagents to cysteines introduced along the length of the pore-lining first TM (TM1) of a cysteine-less variant of CFTR. We find that methanethiosulfonate (MTS) reagents irreversibly modify cysteines substituted for TM1 residues K95, Q98, P99, and L102 when applied to the cytoplasmic side of open channels. Residues closer to the intracellular end of TM1 (Y84-T94) were not apparently modified by MTS reagents, suggesting that this part of TM1 does not line the pore. None of the internal MTS reagent-reactive cysteines was modified by extracellular [2-(trimethylammonium)ethyl] MTS. Only K95C, closest to the putative intracellular end of TM1, was apparently modified by intracellular [2-sulfonatoethyl] MTS before channel activation. Comparison of these results with recent work on CFTR-TM6 suggests a relative alignment of these two important TMs along the axis of the pore. This alignment was tested experimentally by formation of disulfide bridges between pairs of cysteines introduced into these two TMs. Currents carried by the double mutants K95C/I344C and Q98C/I344C, but not by the corresponding single-site mutants, were inhibited by the oxidizing agent copper(II)-o-phenanthroline. This inhibition was irreversible on washing but could be reversed by the reducing agent dithiothreitol, suggesting disulfide bond formation between the introduced cysteine side chains. These results allow us to develop a model of the relative positions, functional contributions, and alignment of two important TMs lining the CFTR pore. Such functional information is necessary to understand and interpret the three-dimensional structure of the pore.
不同的跨膜(TM)α 螺旋已知排列在囊性纤维化跨膜电导调节剂(CFTR)Cl(-)通道的孔中。然而,这些 TM 在孔的三维结构中的相对排列尚不清楚。我们使用膜片钳记录来研究细胞质中应用的半胱氨酸反应性试剂对 CFTR 无半胱氨酸变体的孔衬第一 TM(TM1)中沿长度引入的半胱氨酸的可及性。我们发现,当应用于开放通道的细胞质侧时,甲硫磺酸(MTS)试剂不可逆地修饰取代 TM1 残基 K95、Q98、P99 和 L102 的半胱氨酸。靠近 TM1 细胞内端的残基(Y84-T94)显然没有被 MTS 试剂修饰,这表明这部分 TM1 不排列在孔中。内部 MTS 试剂反应性半胱氨酸中没有一个被细胞外 [2-(三甲基铵)乙基] MTS 修饰。只有靠近 TM1 假设的细胞内端的 K95C,在通道激活之前,显然被细胞内 [2-磺基乙基] MTS 修饰。将这些结果与最近关于 CFTR-TM6 的工作进行比较,表明这两个重要 TM 沿着孔的轴的相对排列。通过将引入这两个 TM 中的成对半胱氨酸形成二硫键来实验测试这种排列。双突变体 K95C/I344C 和 Q98C/I344C 产生的电流,但相应的单突变体没有,被氧化剂铜(II)-邻菲啰啉抑制。这种抑制在洗涤时是不可逆的,但可以被还原剂二硫苏糖醇逆转,这表明引入的半胱氨酸侧链之间形成了二硫键。这些结果使我们能够开发一个模型,用于排列 CFTR 孔衬里的两个重要 TM 的相对位置、功能贡献和排列。这种功能信息对于理解和解释孔的三维结构是必要的。