Department of Medical Protein Research, Ghent University, Ghent, Belgium.
PLoS Genet. 2011 Jul;7(7):e1002169. doi: 10.1371/journal.pgen.1002169. Epub 2011 Jul 7.
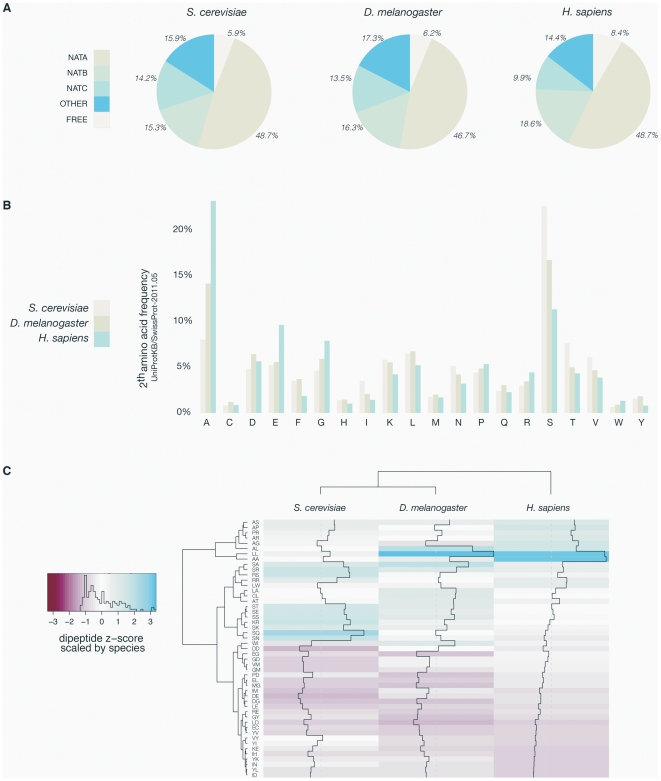
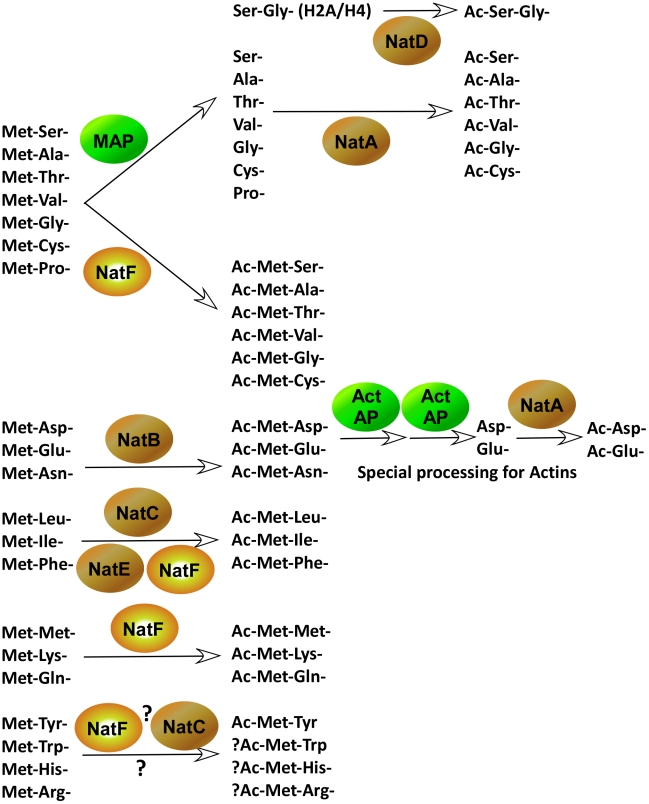
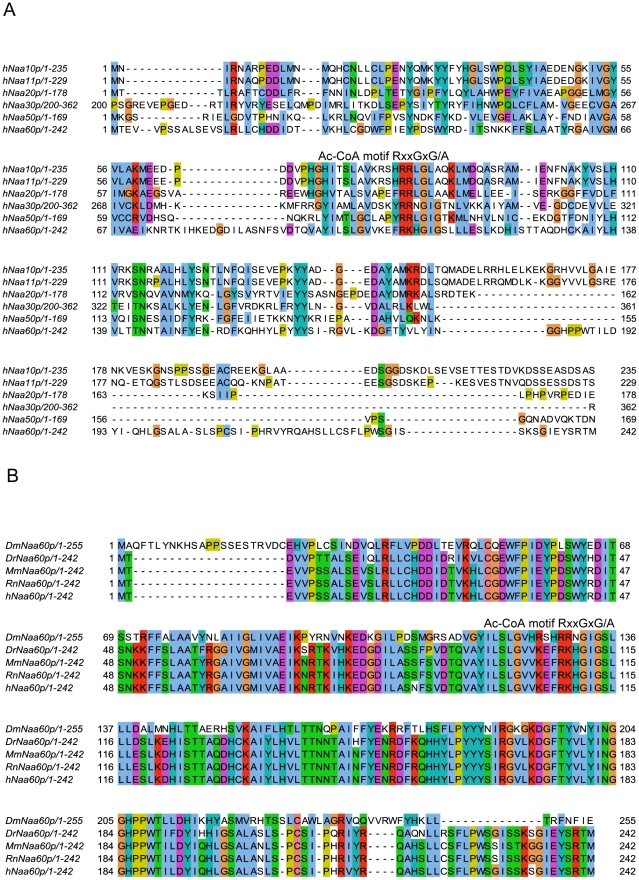
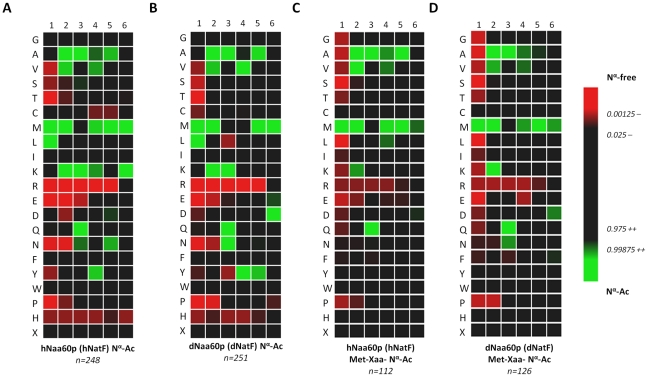
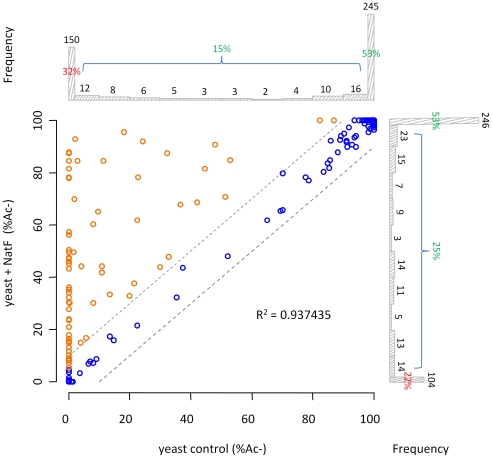
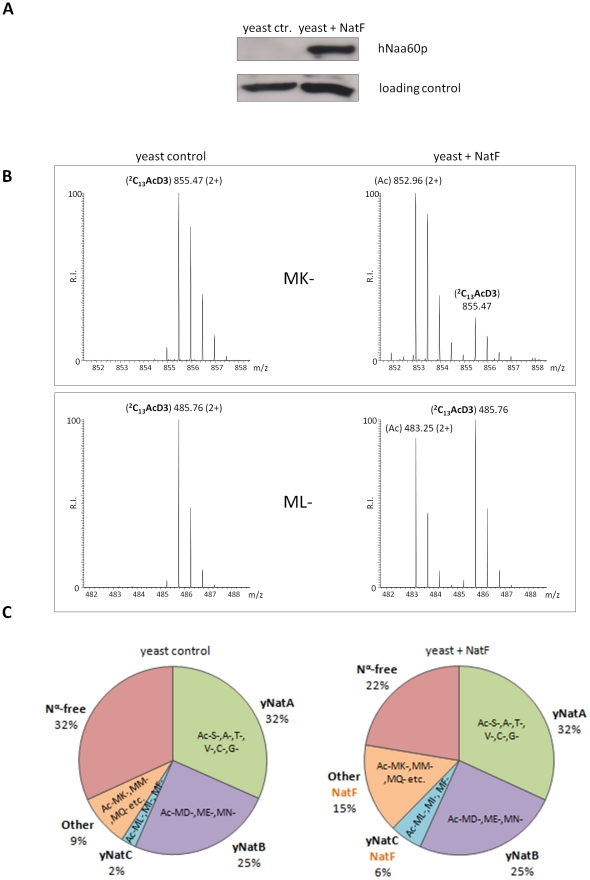
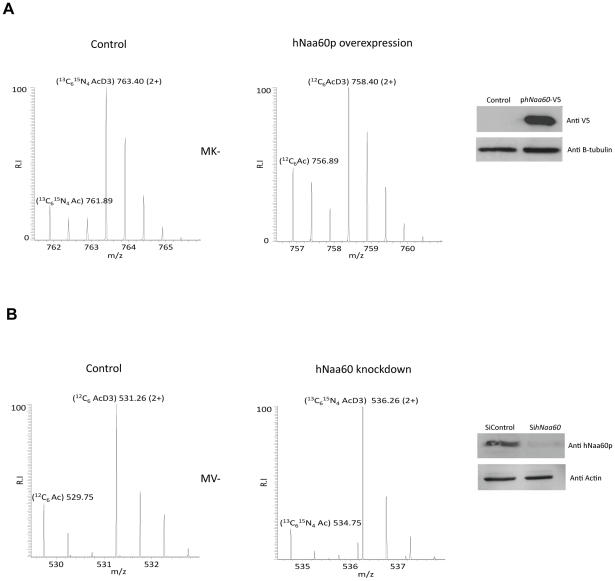
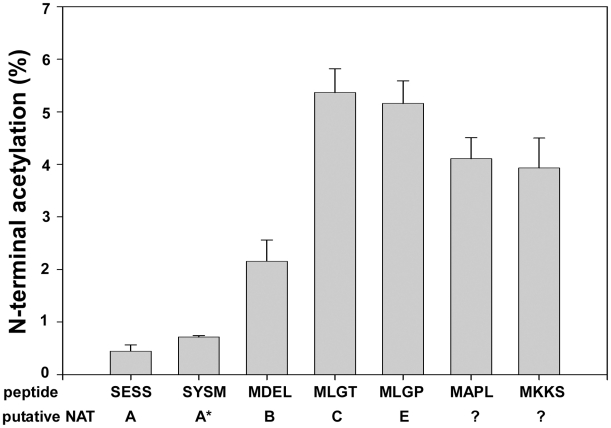
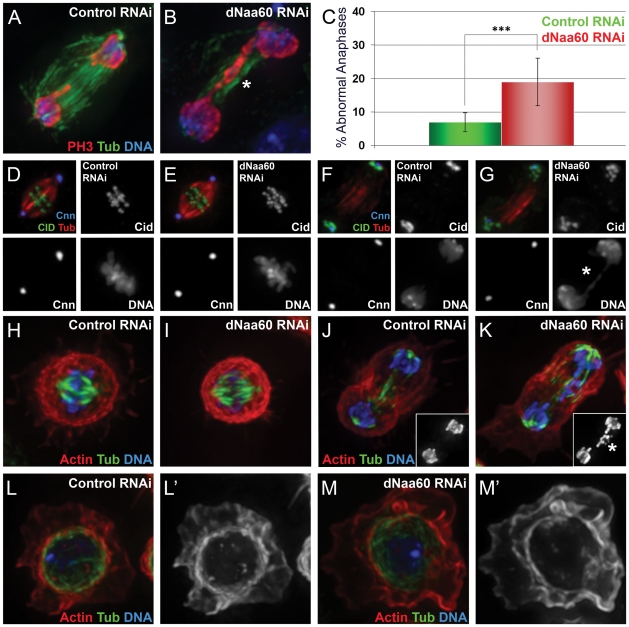
N-terminal acetylation (N-Ac) is a highly abundant eukaryotic protein modification. Proteomics revealed a significant increase in the occurrence of N-Ac from lower to higher eukaryotes, but evidence explaining the underlying molecular mechanism(s) is currently lacking. We first analysed protein N-termini and their acetylation degrees, suggesting that evolution of substrates is not a major cause for the evolutionary shift in N-Ac. Further, we investigated the presence of putative N-terminal acetyltransferases (NATs) in higher eukaryotes. The purified recombinant human and Drosophila homologues of a novel NAT candidate was subjected to in vitro peptide library acetylation assays. This provided evidence for its NAT activity targeting Met-Lys- and other Met-starting protein N-termini, and the enzyme was termed Naa60p and its activity NatF. Its in vivo activity was investigated by ectopically expressing human Naa60p in yeast followed by N-terminal COFRADIC analyses. hNaa60p acetylated distinct Met-starting yeast protein N-termini and increased general acetylation levels, thereby altering yeast in vivo acetylation patterns towards those of higher eukaryotes. Further, its activity in human cells was verified by overexpression and knockdown of hNAA60 followed by N-terminal COFRADIC. NatF's cellular impact was demonstrated in Drosophila cells where NAA60 knockdown induced chromosomal segregation defects. In summary, our study revealed a novel major protein modifier contributing to the evolution of N-Ac, redundancy among NATs, and an essential regulator of normal chromosome segregation. With the characterization of NatF, the co-translational N-Ac machinery appears complete since all the major substrate groups in eukaryotes are accounted for.
N-端乙酰化(N-Ac)是一种高度丰富的真核蛋白修饰。蛋白质组学揭示了从低等到高等真核生物中 N-Ac 的发生率显著增加,但目前缺乏解释潜在分子机制的证据。我们首先分析了蛋白质 N 端及其乙酰化程度,表明底物的进化不是 N-Ac 进化转移的主要原因。此外,我们研究了高等真核生物中潜在的 N-端乙酰转移酶(NAT)的存在。新型 NAT 候选物的纯化重组人源和果蝇同源物进行了体外肽文库乙酰化测定。这为其靶向 Met-Lys-和其他 Met 起始蛋白 N 端的 NAT 活性提供了证据,该酶被命名为 Naa60p 和其活性 NatF。通过在酵母中异位表达人源 Naa60p 然后进行 N 端 COFRADIC 分析来研究其体内活性。hNaa60p 乙酰化了不同的 Met 起始酵母蛋白 N 端,并增加了总体乙酰化水平,从而改变了酵母体内的乙酰化模式,使其向高等真核生物的模式靠拢。此外,通过人源 NAA60 的过表达和敲低,然后进行 N 端 COFRADIC,验证了其在人细胞中的活性。在果蝇细胞中证实了 NatF 的细胞作用,其中 NAA60 敲低诱导了染色体分离缺陷。总之,我们的研究揭示了一种新的主要蛋白质修饰物,它有助于 N-Ac 的进化、NAT 之间的冗余性以及正常染色体分离的重要调节因子。随着 NatF 的特性描述,共翻译 N-Ac 机制似乎已经完整,因为真核生物中的所有主要底物群都得到了说明。