Plant Development and Nuclear Architecture, Centro de Investigaciones Biológicas, Madrid, Spain.
J Exp Bot. 2012 Mar;63(5):2007-24. doi: 10.1093/jxb/err400. Epub 2011 Dec 23.
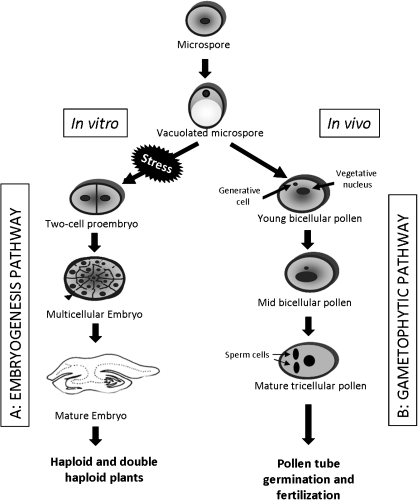
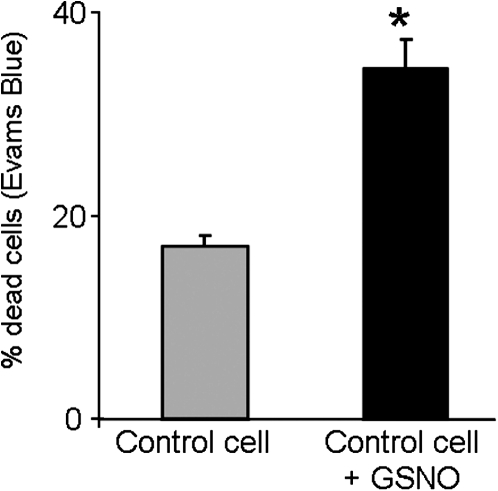
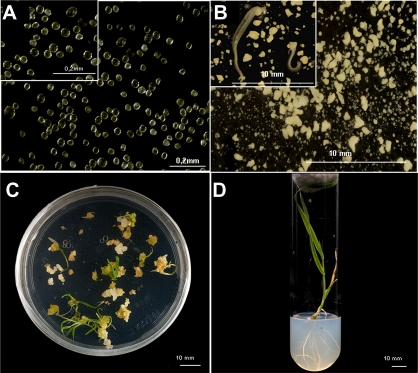
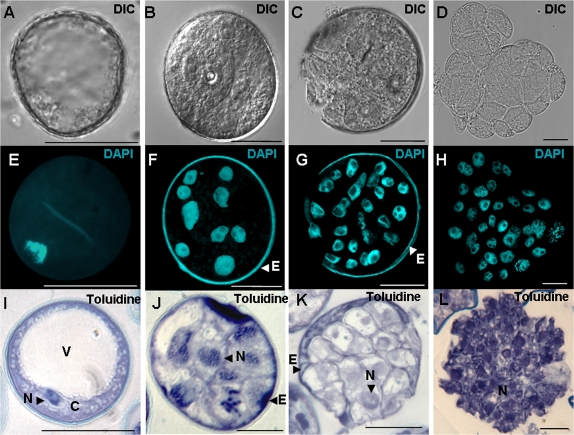
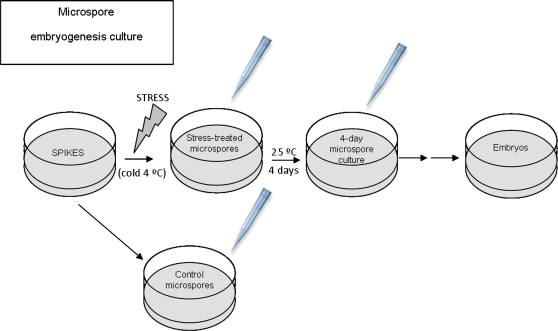
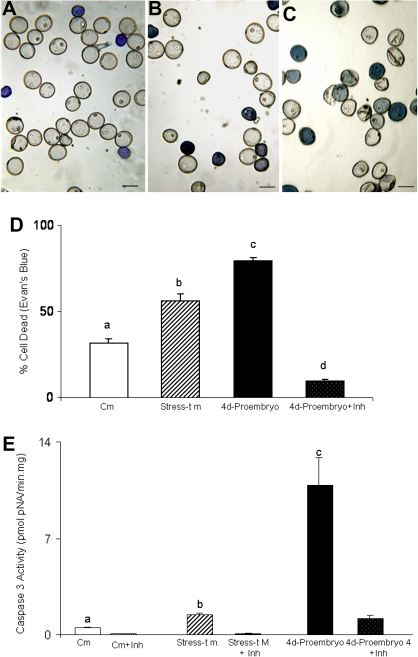
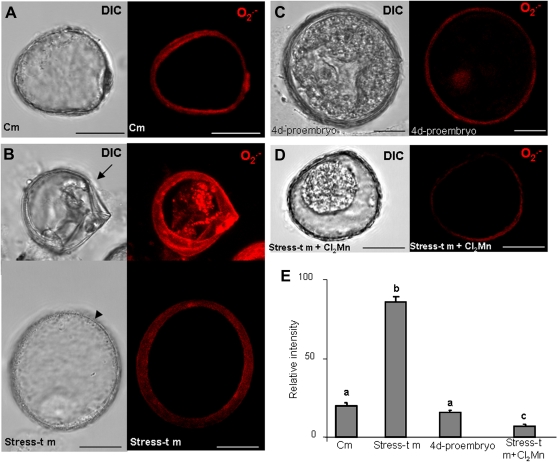
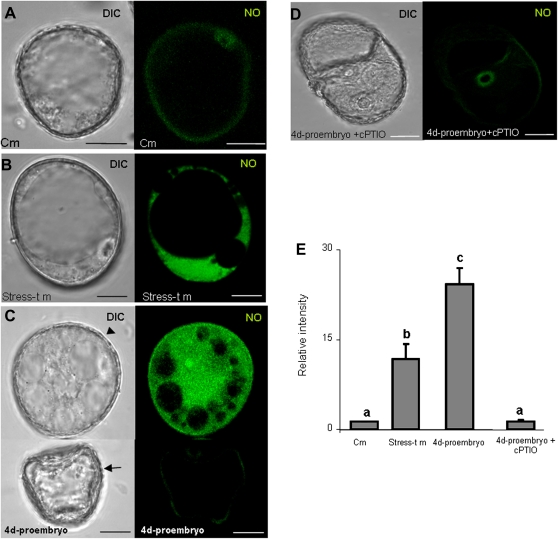
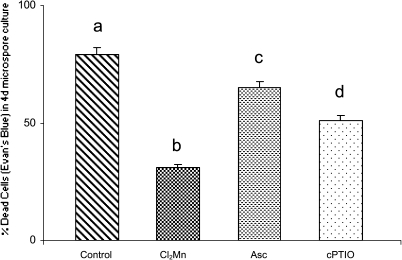
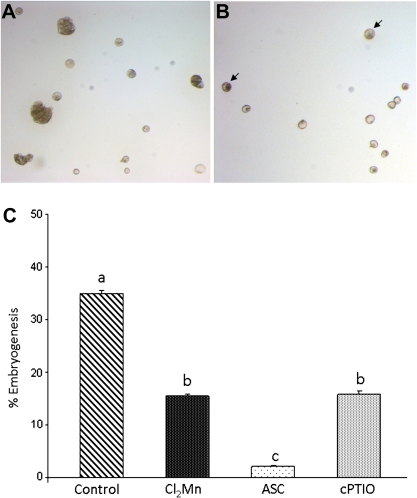
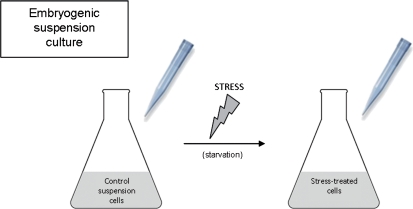
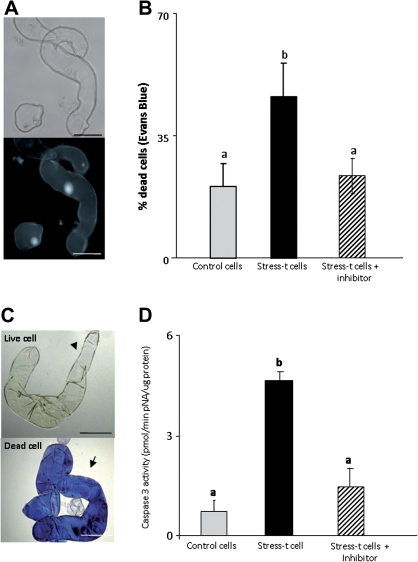
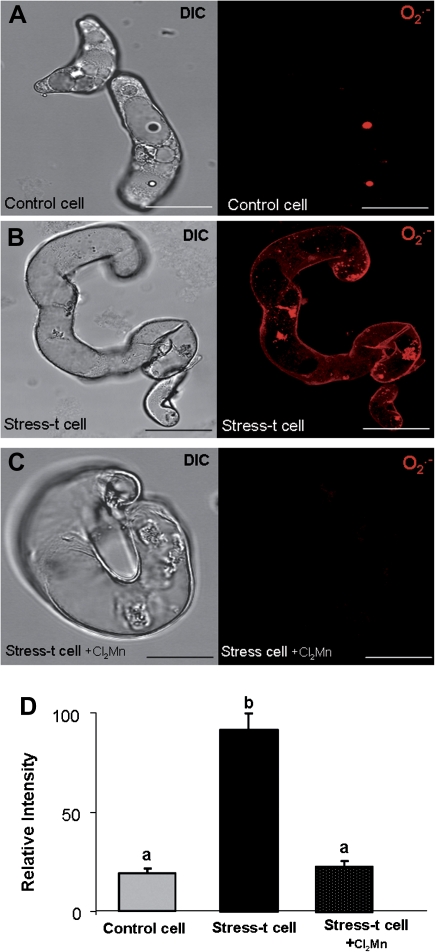
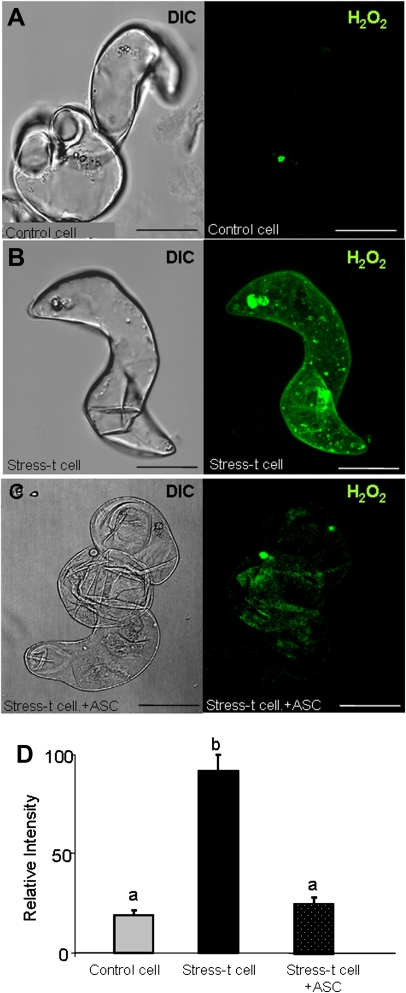
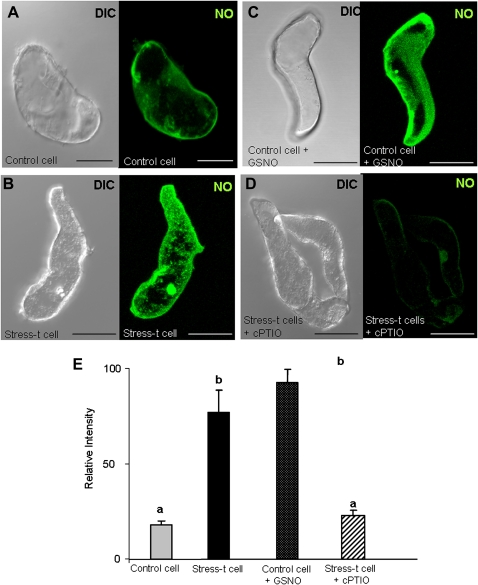
Under specific stress treatments (cold, starvation), in vitro microspores can be induced to deviate from their gametophytic development and switch to embryogenesis, forming haploid embryos and homozygous breeding lines in a short period of time. The inductive stress produces reactive oxygen species (ROS) and nitric oxide (NO), signalling molecules mediating cellular responses, and cell death, modifying the embryogenic microspore response and therefore, the efficiency of the process. This work analysed cell death, caspase 3-like activity, and ROS and NO production (using fluorescence probes and confocal analysis) after inductive stress in barley microspore cultures and embryogenic suspension cultures, as an in vitro system which permitted easy handling for comparison. There was an increase in caspase 3-like activity and cell death after stress treatment in microspore and suspension cultures, while ROS increased in non-induced microspores and suspension cultures. Treatments of the cultures with a caspase 3 inhibitor, DEVD-CHO, significantly reduced the cell death percentages. Stress-treated embryogenic suspension cultures exhibited high NO signals and cell death, while treatment with S-nitrosoglutathione (NO donor) in control suspension cultures resulted in even higher cell death. In contrast, in microspore cultures, NO production was detected after stress, and, in the case of 4-day microspore cultures, in embryogenic microspores accompanying the initiation of cell divisions. Subsequent treatments of stress-treated microspore cultures with ROS and NO scavengers resulted in a decreasing cell death during the early stages, but later they produced a delay in embryo development as well as a decrease in the percentage of embryogenesis in microspores. Results showed that the ROS increase was involved in the stress-induced programmed cell death occurring at early stages in both non-induced microspores and embryogenic suspension cultures; whereas NO played a dual role after stress in the two in vitro systems, one involved in programmed cell death in embryogenic suspension cultures and the other in the initiation of cell division leading to embryogenesis in reprogrammed microspores.
在特定的胁迫处理(冷、饥饿)下,体外小孢子可以偏离其配子体发育并转向胚胎发生,在短时间内形成单倍体胚胎和纯合系。诱导胁迫会产生活性氧(ROS)和一氧化氮(NO),作为信号分子介导细胞反应和细胞死亡,改变胚胎发生小孢子的反应,从而改变该过程的效率。本研究分析了在大麦小孢子培养物和胚胎发生悬浮培养物中,诱导胁迫后细胞死亡、半胱天冬酶 3 样活性以及 ROS 和 NO 产生(使用荧光探针和共聚焦分析),作为一个易于处理的体外系统进行比较。在小孢子和悬浮培养物中,胁迫处理后半胱天冬酶 3 样活性和细胞死亡增加,而 ROS 在未诱导的小孢子和悬浮培养物中增加。用半胱天冬酶 3 抑制剂 DEVD-CHO 处理培养物可显著降低细胞死亡百分比。胁迫处理的胚胎发生悬浮培养物表现出高的 NO 信号和细胞死亡,而用 S-亚硝基谷胱甘肽(NO 供体)处理对照悬浮培养物则导致更高的细胞死亡。相反,在小孢子培养物中,在胁迫后检测到 NO 产生,在 4 天小孢子培养物中,在胚胎发生小孢子中伴随细胞分裂的开始。随后用 ROS 和 NO 清除剂处理胁迫处理的小孢子培养物,在早期阶段会降低细胞死亡,但后来会延迟胚胎发育,并降低小孢子的胚胎发生百分比。结果表明,ROS 增加参与了在非诱导小孢子和胚胎发生悬浮培养物早期发生的胁迫诱导的程序性细胞死亡;而 NO 在两个体外系统中的胁迫后起双重作用,一种涉及胚胎发生悬浮培养物中的程序性细胞死亡,另一种涉及导致重新编程小孢子胚胎发生的细胞分裂的开始。