Computational Neuroscience Group, Department of Integrative Neurophysiology, Center for Neurogenomics and Cognitive Research, VU University Amsterdam Amsterdam, Netherlands.
Front Comput Neurosci. 2012 Jan 25;5:54. doi: 10.3389/fncom.2011.00054. eCollection 2011.
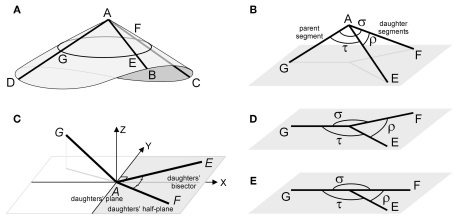
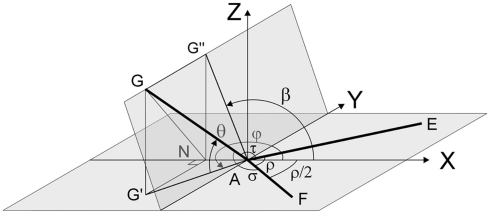
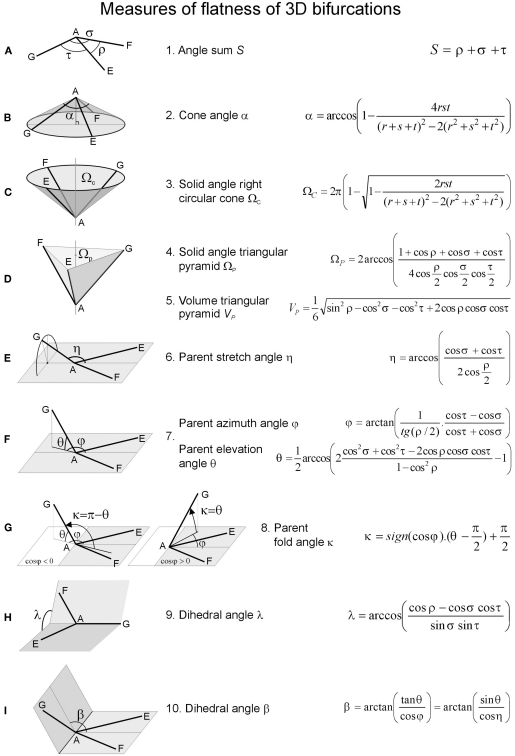
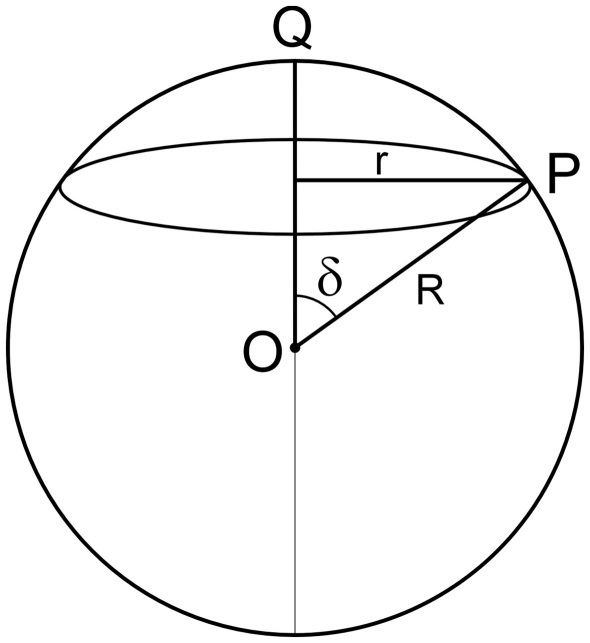
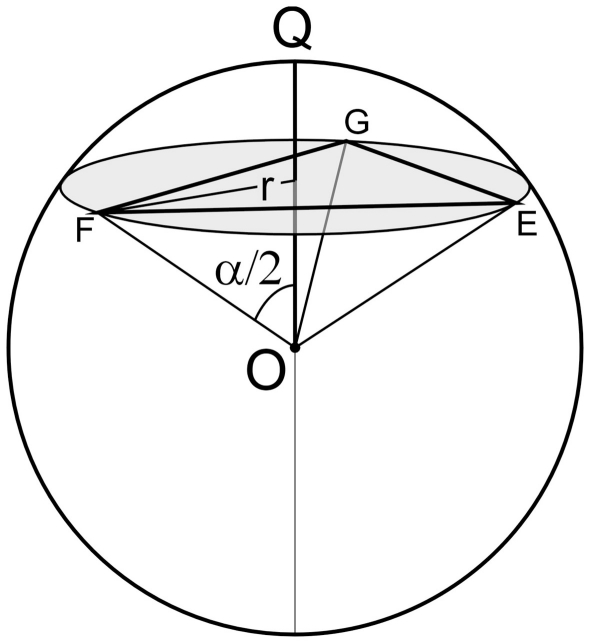
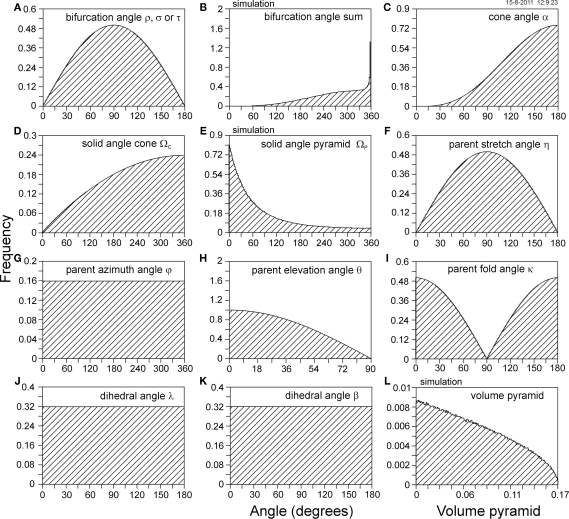
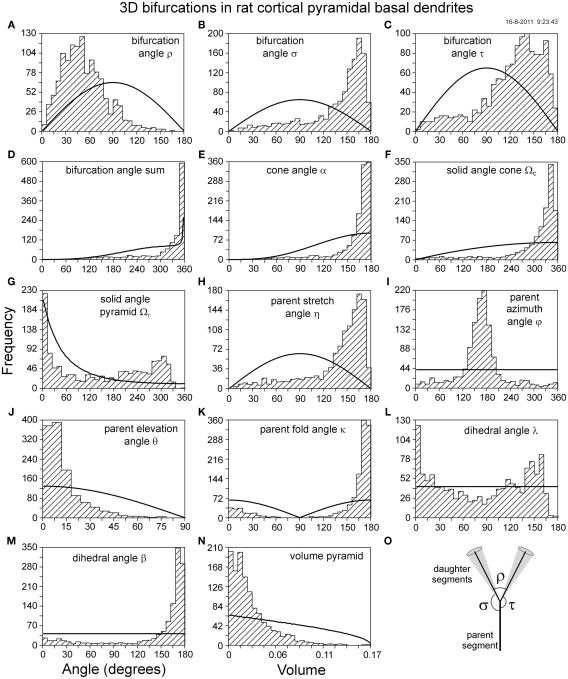
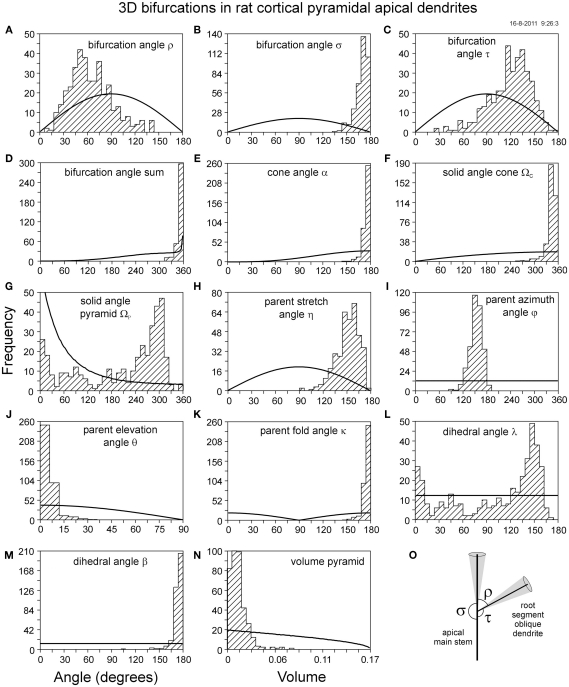
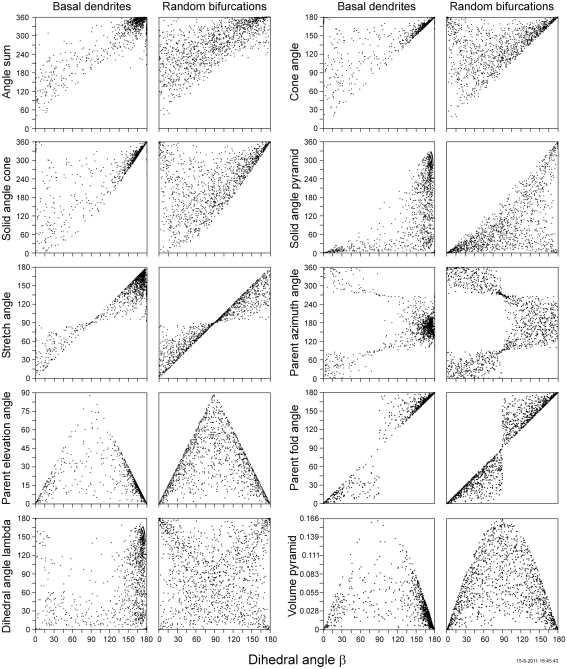
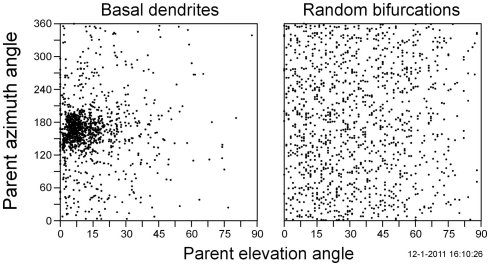
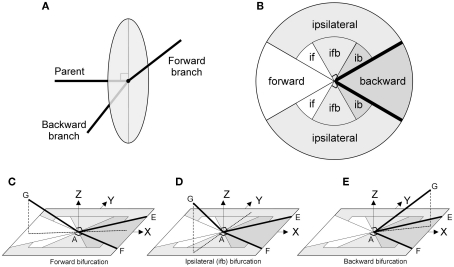
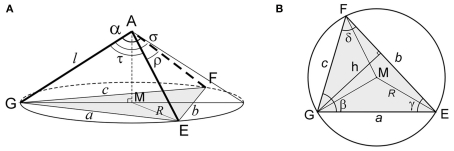
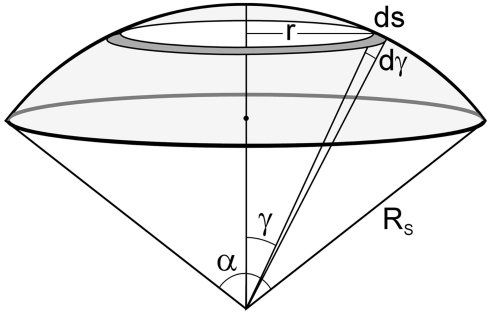
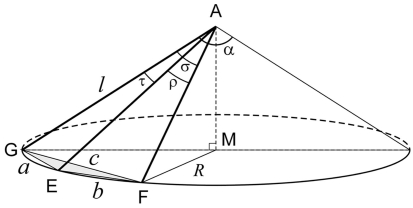
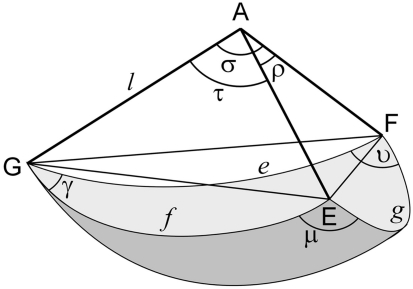
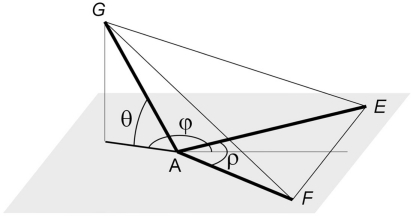
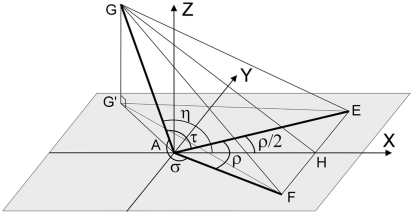
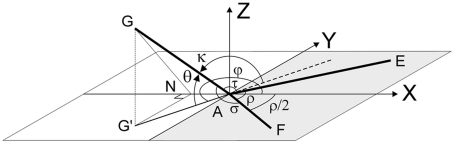
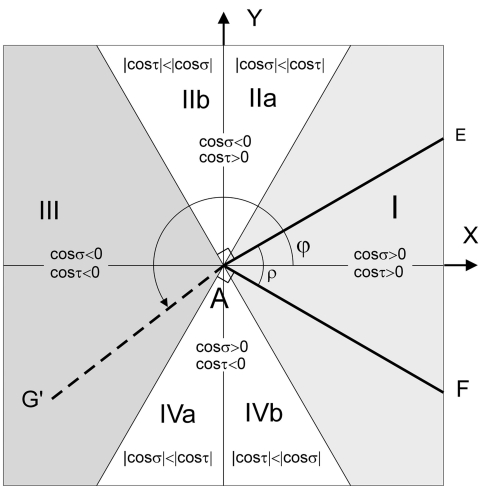
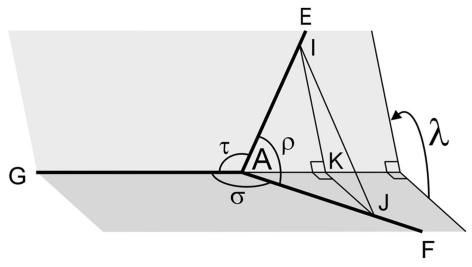
The geometry of natural branching systems generally reflects functional optimization. A common property is that their bifurcations are planar and that daughter segments do not turn back in the direction of the parent segment. The present study investigates whether this also applies to bifurcations in 3D dendritic arborizations. This question was earlier addressed in a first study of flatness of 3D dendritic bifurcations by Uylings and Smit (1975), who used the apex angle of the right circular cone as flatness measure. The present study was inspired by recent renewed interest in this measure. Because we encountered ourselves shortcomings of this cone angle measure, the search for an optimal measure for flatness of 3D bifurcation was the second aim of our study. Therefore, a number of measures has been developed in order to quantify flatness and orientation properties of spatial bifurcations. All these measures have been expressed mathematically in terms of the three bifurcation angles between the three pairs of segments in the bifurcation. The flatness measures have been applied and evaluated to bifurcations in rat cortical pyramidal cell basal and apical dendritic trees, and to random spatial bifurcations. Dendritic and random bifurcations show significant different flatness measure distributions, supporting the conclusion that dendritic bifurcations are significantly more flat than random bifurcations. Basal dendritic bifurcations also show the property that their parent segments are generally aligned oppositely to the bisector of the angle between their daughter segments, resulting in "symmetrical" configurations. Such geometries may arise when during neuronal development the segments at a newly formed bifurcation are subjected to elastic tensions, which force the bifurcation into an equilibrium planar shape. Apical bifurcations, however, have parent segments oppositely aligned with one of the daughter segments. These geometries arise in the case of side branching from an existing apical main stem. The aligned "apical" parent and "apical" daughter segment form together with the side branch daughter segment already geometrically a flat configuration. These properties are clearly reflected in the flatness measure distributions. Comparison of the different flatness measures made clear that they all capture flatness properties in a different way. Selection of the most appropriate measure thus depends on the question of research. For our purpose of quantifying flatness and orientation of the segments, the dihedral angle β was found to be the most discriminative and applicable single measure. Alternatively, the parent elevation and azimuth angle formed an orthogonal pair of measures most clearly demonstrating the dendritic bifurcation "symmetry" properties.
自然分支系统的几何形状通常反映了功能优化。一个共同的特点是它们的分支是平面的,并且子段不会朝着父段的方向返回。本研究探讨了这是否也适用于 3D 树突分支的分支。Uylings 和 Smit(1975)最早在对 3D 树突分支的平坦度的第一项研究中探讨了这个问题,他们使用右圆锥的顶点角作为平坦度的度量。本研究的灵感来自于最近对该度量的重新关注。由于我们发现这个圆锥角度量存在缺陷,因此寻找 3D 分支平坦度的最佳度量是我们研究的第二个目标。因此,为了量化空间分支的平坦度和方向特性,开发了许多度量。所有这些度量都以分支中三个分支对之间的三个分支角的形式在数学上表示。这些平坦度度量已经应用于并评估了大鼠皮质锥体细胞基底和顶树突分支的分支,以及随机空间分支。树突和随机分支显示出显著不同的平坦度度量分布,支持树突分支比随机分支更平坦的结论。基底树突分支还表现出它们的父段通常与它们的子段之间的夹角的平分线相反对齐的性质,从而导致“对称”的构型。当新形成的分支处的段受到弹性张力时,可能会出现这种几何形状,这些张力迫使分支进入平衡的平面形状。然而,顶树突分支的父段与一个子段相反对齐。这些几何形状出现在现有顶主干的侧分支的情况下。对齐的“顶”父段和“顶”子段与侧分支子段一起已经形成了一个平坦的构型。这些特性在平坦度度量分布中得到了清晰的反映。不同平坦度度量的比较清楚地表明,它们以不同的方式捕捉平坦度特性。因此,最合适的度量的选择取决于研究的问题。对于我们量化段的平坦度和方向的目的,二面角 β 被发现是最具区分性和适用的单一度量。或者,父段的仰角和方位角形成了一对最能清楚地展示树突分支“对称”特性的正交度量。