Hubei Key Laboratory of Genetic Regulation and Integrative Biology, College of Life Science, Huazhong Normal University, Wuhan, People's Republic of China.
PLoS One. 2012;7(2):e30375. doi: 10.1371/journal.pone.0030375. Epub 2012 Feb 17.
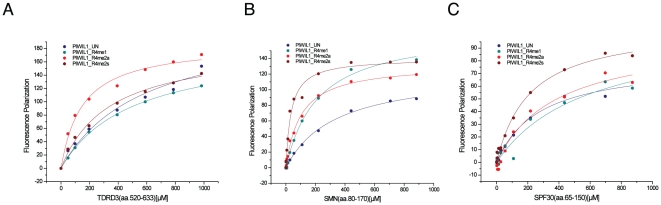
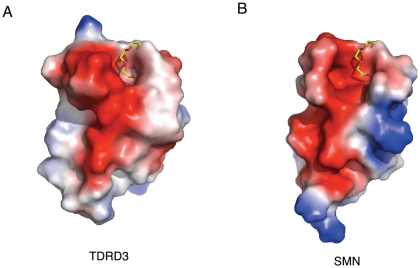
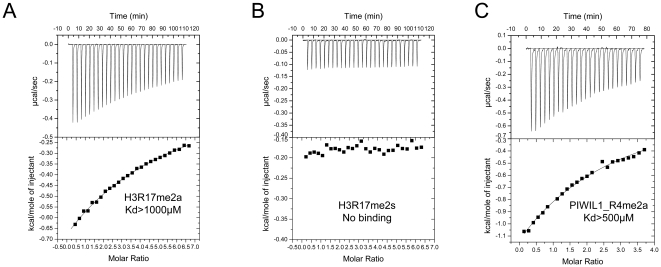
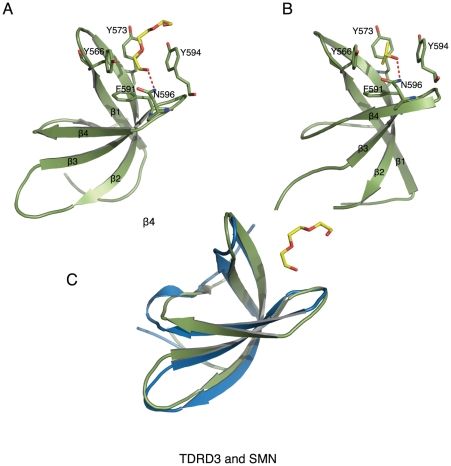
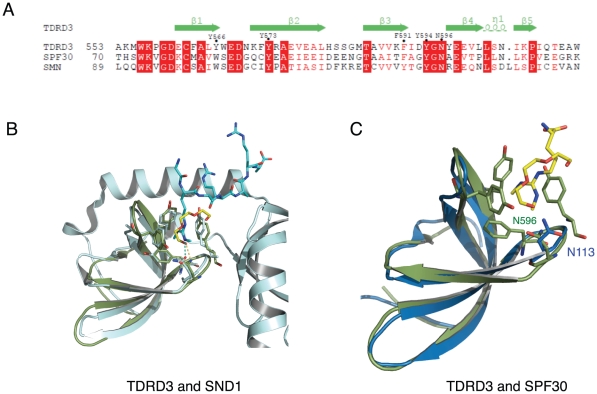
SMN (Survival motor neuron protein) was characterized as a dimethyl-arginine binding protein over ten years ago. TDRD3 (Tudor domain-containing protein 3) and SPF30 (Splicing factor 30 kDa) were found to bind to various methyl-arginine proteins including Sm proteins as well later on. Recently, TDRD3 was shown to be a transcriptional coactivator, and its transcriptional activity is dependent on its ability to bind arginine-methylated histone marks. In this study, we systematically characterized the binding specificity and affinity of the Tudor domains of these three proteins quantitatively. Our results show that TDRD3 preferentially recognizes asymmetrical dimethylated arginine mark, and SMN is a very promiscuous effector molecule, which recognizes different arginine containing sequence motifs and preferentially binds symmetrical dimethylated arginine. SPF30 is the weakest methyl-arginine binder, which only binds the GAR motif sequences in our library. In addition, we also reported high-resolution crystal structures of the Tudor domain of TDRD3 in complex with two small molecules, which occupy the aromatic cage of TDRD3.
SMN(运动神经元存活蛋白)早在十年前就被确定为一种二甲基精氨酸结合蛋白。TDRD3(含 Tudor 结构域蛋白 3)和 SPF30(30kDa 剪接因子)后来也被发现与包括 Sm 蛋白在内的各种甲基精氨酸蛋白结合。最近,TDRD3 被证明是一种转录共激活因子,其转录活性依赖于其结合精氨酸甲基化组蛋白标记的能力。在这项研究中,我们系统地定量表征了这三种蛋白质的 Tudor 结构域的结合特异性和亲和力。我们的结果表明,TDRD3 优先识别非对称二甲基精氨酸标记,而 SMN 是一种非常混杂的效应分子,它识别不同的精氨酸含有序列基序,并优先结合对称二甲基精氨酸。SPF30 是最弱的精氨酸结合物,仅在我们的文库中结合 GAR 基序序列。此外,我们还报道了 TDRD3 的 Tudor 结构域与两种小分子复合物的高分辨率晶体结构,这两种小分子占据了 TDRD3 的芳香笼。