Computational & Systems Biology, John Innes Centre, Norwich Research Park, Norwich, United Kingdom.
PLoS Comput Biol. 2012;8(3):e1002402. doi: 10.1371/journal.pcbi.1002402. Epub 2012 Mar 1.
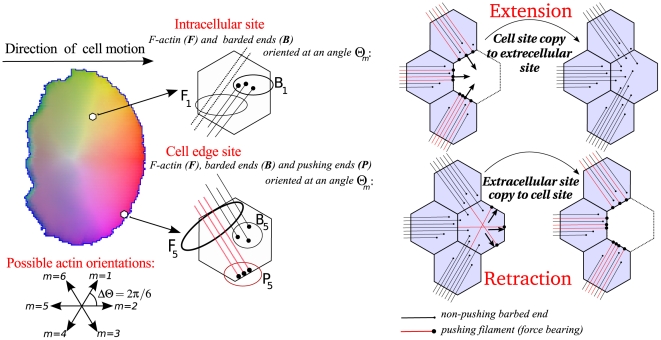
To regulate shape changes, motility and chemotaxis in eukaryotic cells, signal transduction pathways channel extracellular stimuli to the reorganization of the actin cytoskeleton. The complexity of such networks makes it difficult to understand the roles of individual components, let alone their interactions and multiple feedbacks within a given layer and between layers of signalling. Even more challenging is the question of if and how the shape of the cell affects and is affected by this internal spatiotemporal reorganization. Here we build on our previous 2D cell motility model where signalling from the Rho family GTPases (Cdc42, Rac, and Rho) was shown to organize the cell polarization, actin reorganization, shape change, and motility in simple gradients. We extend this work in two ways: First, we investigate the effects of the feedback between the phosphoinositides (PIs) PIP₂, PIP₃ and Rho family GTPases. We show how that feedback increases heights and breadths of zones of Cdc42 activity, facilitating global communication between competing cell "fronts". This hastens the commitment to a single lamellipodium initiated in response to multiple, complex, or rapidly changing stimuli. Second, we show how cell shape feeds back on internal distribution of GTPases. Constraints on chemical isocline curvature imposed by boundary conditions results in the fact that dynamic cell shape leads to faster biochemical redistribution when the cell is repolarized. Cells with frozen cytoskeleton, and static shapes, consequently respond more slowly to reorienting stimuli than cells with dynamic shape changes, the degree of the shape-induced effects being proportional to the extent of cell deformation. We explain these concepts in the context of several in silico experiments using our 2D computational cell model.
为了调节真核细胞的形状变化、运动和趋化性,信号转导途径将细胞外刺激传递到肌动蛋白细胞骨架的重排。这些网络的复杂性使得难以理解单个成分的作用,更不用说它们在给定信号层内以及在不同信号层之间的相互作用和多重反馈了。更具挑战性的问题是,细胞的形状是否以及如何影响并受到这种内部时空重排的影响。在这里,我们基于我们之前的 2D 细胞运动模型,该模型表明 Rho 家族 GTPases(Cdc42、Rac 和 Rho)的信号转导组织了细胞极化、肌动蛋白重排、形状变化和简单梯度中的运动。我们以两种方式扩展了这项工作:首先,我们研究了磷酸肌醇(PIs)PIP₂、PIP₃ 和 Rho 家族 GTPases 之间的反馈的影响。我们展示了这种反馈如何增加 Cdc42 活性区的高度和宽度,促进竞争细胞“前沿”之间的全局通讯。这加速了对多个复杂或快速变化的刺激做出单一片状伪足启动的承诺。其次,我们展示了细胞形状如何反馈内部 GTPase 分布。边界条件对化学等变线曲率的约束导致动态细胞形状导致在细胞重新极化时更快的生化再分配。与具有动态形状变化的细胞相比,具有冻结的细胞骨架和静态形状的细胞对重新定向刺激的反应更慢,形状引起的效应的程度与细胞变形的程度成正比。我们使用我们的 2D 计算细胞模型在几个模拟实验中解释了这些概念。