Department of Biochemistry and Molecular Biology, University of Texas MD Anderson Cancer Center, Houston, TX, 77030, USA.
Department of Pharmacology and Structural Biology and Biophysics Program, University of Colorado, School of Medicine, 12801 East 17th Avenue, Aurora, CO, 80045-0511, USA.
Epigenetics Chromatin. 2012 Apr 27;5(1):5. doi: 10.1186/1756-8935-5-5.
The protein anti-silencing function 1 (Asf1) chaperones histones H3/H4 for assembly into nucleosomes every cell cycle as well as during DNA transcription and repair. Asf1 interacts directly with H4 through the C-terminal tail of H4, which itself interacts with the docking domain of H2A in the nucleosome. The structure of this region of the H4 C-terminus differs greatly in these two contexts.
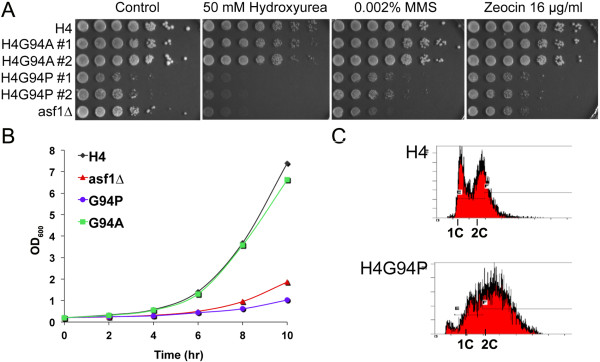
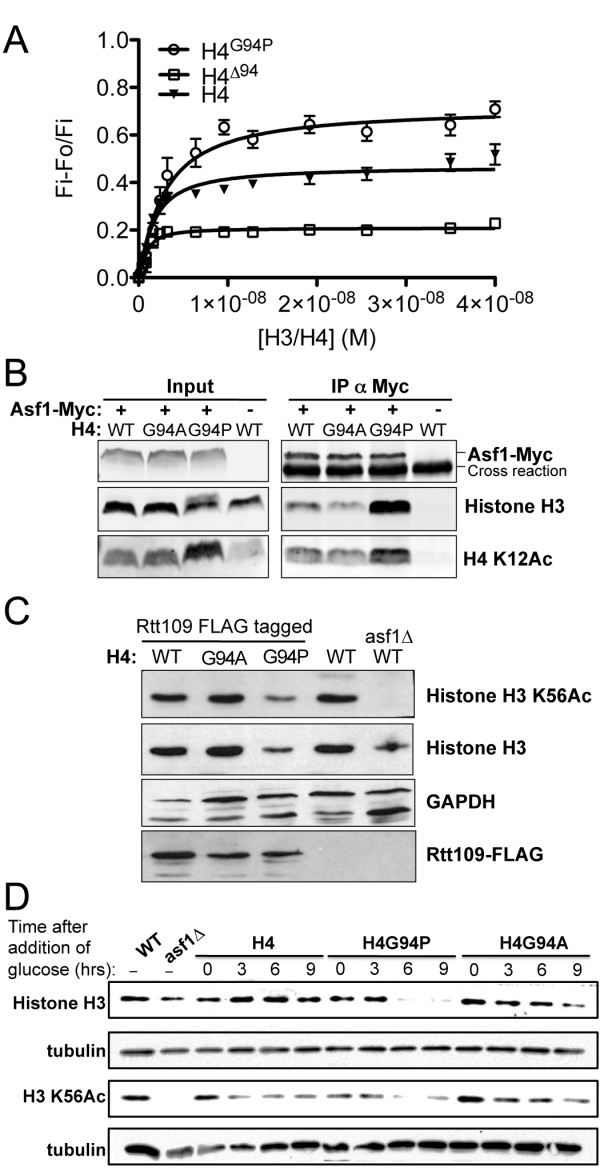
To investigate the functional consequence of this structural change in histone H4, we restricted the available conformations of the H4 C-terminus and analyzed its effect in vitro and in vivo in Saccharomyces cerevisiae. One such mutation, H4 G94P, had modest effects on the interaction between H4 and Asf1. However, in yeast, flexibility of the C-terminal tail of H4 has essential functions that extend beyond chromatin assembly and disassembly. The H4 G94P mutation resulted in severely sick yeast, although nucleosomes still formed in vivo albeit yielding diffuse micrococcal nuclease ladders. In vitro, H4G4P had modest effects on nucleosome stability, dramatically reduced histone octamer stability, and altered nucleosome sliding ability.
The functional consequences of altering the conformational flexibility in the C-terminal tail of H4 are severe. Interestingly, despite the detrimental effects of the histone H4 G94P mutant on viability, nucleosome formation was not markedly affected in vivo. However, histone octamer stability and nucleosome stability as well as nucleosome sliding ability were altered in vitro. These studies highlight an important role for correct interactions of the histone H4 C-terminal tail within the histone octamer and suggest that maintenance of a stable histone octamer in vivo is an essential feature of chromatin dynamics.
蛋白质抗沉默功能 1(Asf1)作为伴侣蛋白,在每个细胞周期中以及在 DNA 转录和修复过程中,将组蛋白 H3/H4 组装成核小体。Asf1 通过 H4 的 C 末端与 H4 直接相互作用,而 H4 本身与核小体中 H2A 的对接域相互作用。在这两种情况下,H4 C 末端的这一区域的结构有很大的不同。
为了研究组蛋白 H4 中这种结构变化的功能后果,我们限制了 H4 C 末端的可用构象,并在体外和酿酒酵母体内分析了其影响。这样的突变之一,H4 G94P,对 H4 和 Asf1 之间的相互作用有轻微影响。然而,在酵母中,H4 C 末端的灵活性具有超出染色质组装和拆卸的重要功能。H4 G94P 突变导致酵母严重患病,尽管体内仍能形成核小体,但产生弥散的微球菌核酸酶梯。在体外,H4G4P 对核小体稳定性有轻微影响,显著降低了组蛋白八聚体稳定性,并改变了核小体滑动能力。
改变 H4 C 末端构象灵活性的功能后果是严重的。有趣的是,尽管组蛋白 H4 G94P 突变对生存能力有不利影响,但体内核小体的形成并没有明显受到影响。然而,组蛋白八聚体稳定性、核小体稳定性以及核小体滑动能力在体外发生了改变。这些研究强调了正确的组蛋白 H4 C 末端在组蛋白八聚体中的相互作用的重要作用,并表明在体内维持稳定的组蛋白八聚体是染色质动力学的一个重要特征。