Sanford-Burnham Medical Research Institute, La Jolla, California, United States of America.
PLoS One. 2012;7(6):e39115. doi: 10.1371/journal.pone.0039115. Epub 2012 Jun 15.
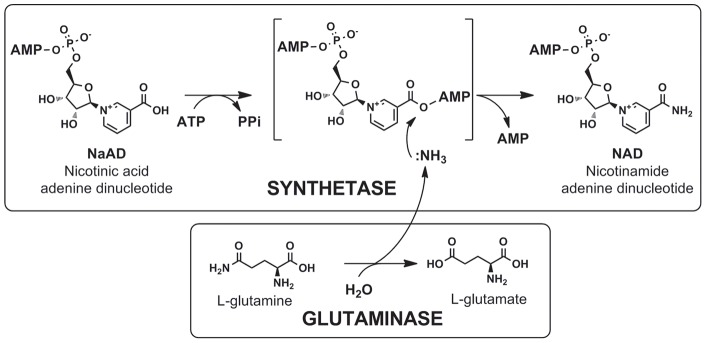
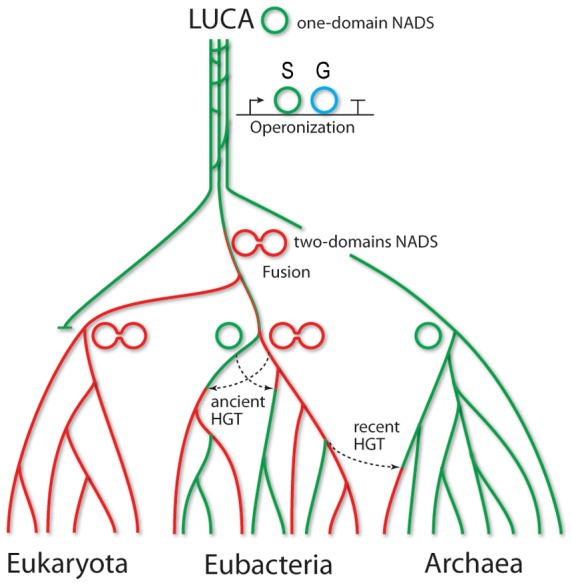
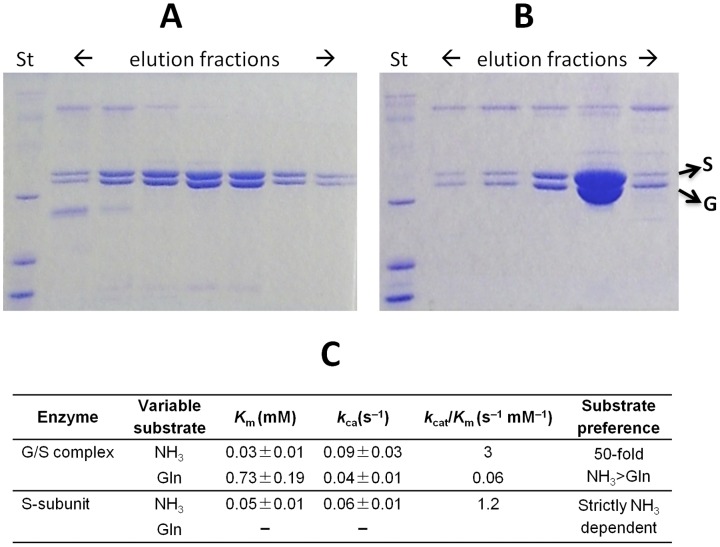
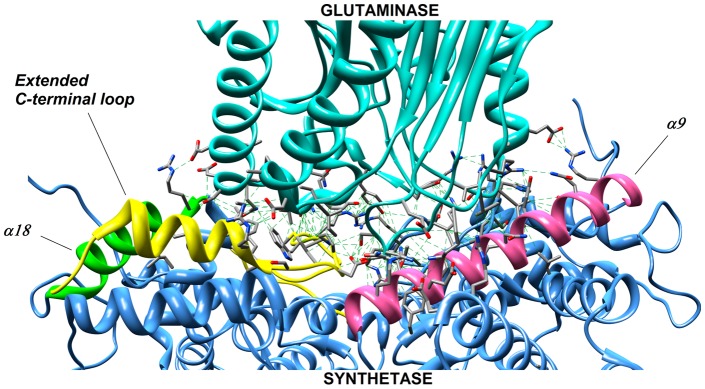
NAD is a ubiquitous and essential metabolic redox cofactor which also functions as a substrate in certain regulatory pathways. The last step of NAD synthesis is the ATP-dependent amidation of deamido-NAD by NAD synthetase (NADS). Members of the NADS family are present in nearly all species across the three kingdoms of Life. In eukaryotic NADS, the core synthetase domain is fused with a nitrilase-like glutaminase domain supplying ammonia for the reaction. This two-domain NADS arrangement enabling the utilization of glutamine as nitrogen donor is also present in various bacterial lineages. However, many other bacterial members of NADS family do not contain a glutaminase domain, and they can utilize only ammonia (but not glutamine) in vitro. A single-domain NADS is also characteristic for nearly all Archaea, and its dependence on ammonia was demonstrated here for the representative enzyme from Methanocaldococcus jannaschi. However, a question about the actual in vivo nitrogen donor for single-domain members of the NADS family remained open: Is it glutamine hydrolyzed by a committed (but yet unknown) glutaminase subunit, as in most ATP-dependent amidotransferases, or free ammonia as in glutamine synthetase? Here we addressed this dilemma by combining evolutionary analysis of the NADS family with experimental characterization of two representative bacterial systems: a two-subunit NADS from Thermus thermophilus and a single-domain NADS from Salmonella typhimurium providing evidence that ammonia (and not glutamine) is the physiological substrate of a typical single-domain NADS. The latter represents the most likely ancestral form of NADS. The ability to utilize glutamine appears to have evolved via recruitment of a glutaminase subunit followed by domain fusion in an early branch of Bacteria. Further evolution of the NADS family included lineage-specific loss of one of the two alternative forms and horizontal gene transfer events. Lastly, we identified NADS structural elements associated with glutamine-utilizing capabilities.
NAD 是一种普遍存在且必需的代谢氧化还原辅助因子,也是某些调节途径中的底物。NAD 合成的最后一步是 NAD 合成酶(NADS)依赖于 ATP 的将去酰胺 NAD 酰胺化。NADS 家族的成员存在于生命的三个王国中的几乎所有物种中。在真核生物 NADS 中,核心合成酶结构域与类似腈酶的谷氨酰胺酶结构域融合,为反应提供氨。这种具有两个结构域的 NADS 排列方式使 glutamine 能够作为氮供体被利用,这种方式也存在于各种细菌谱系中。然而,NADS 家族的许多其他细菌成员不含谷氨酰胺酶结构域,它们只能在体外利用氨(而不是 glutamine)。单个结构域 NADS 也是几乎所有古菌的特征,这里证明了来自 Methanocaldococcus jannaschi 的代表性酶对氨的依赖性。然而,关于 NADS 家族单个结构域成员的实际体内氮供体的问题仍然存在:它是像大多数 ATP 依赖性氨转移酶那样,由一个特定(但尚未确定)的谷氨酰胺酶亚基水解 glutamine 产生,还是像 glutamine 合酶那样由游离氨产生?在这里,我们通过对 NADS 家族的进化分析与对两个代表性细菌系统(来自 Thermus thermophilus 的二亚基 NADS 和来自 Salmonella typhimurium 的单个结构域 NADS)的实验特性分析相结合,解决了这个难题,为氨(而非 glutamine)是典型的单个结构域 NADS 的生理底物提供了证据。后者代表了最有可能的 NADS 原始形式。利用 glutamine 的能力似乎是通过在细菌的早期分支中招募一个谷氨酰胺酶亚基并随后进行结构域融合而进化而来的。NADS 家族的进一步进化包括两种替代形式中的一种在特定谱系中的丢失和水平基因转移事件。最后,我们确定了与利用 glutamine 能力相关的 NADS 结构元素。