Department of Computational and Systems Biology, School of Medicine, University of Pittsburgh, Pittsburgh, Pennsylvania 15213, USA.
J Biol Chem. 2012 Sep 14;287(38):31823-32. doi: 10.1074/jbc.M112.397364. Epub 2012 Jul 29.
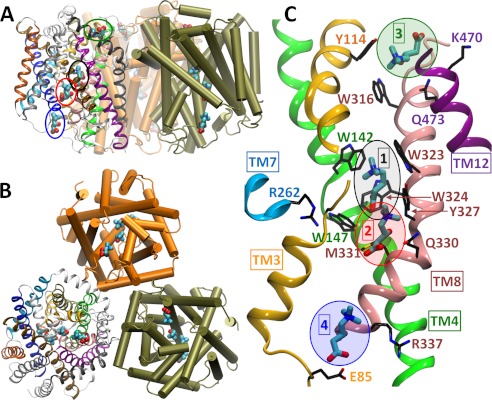
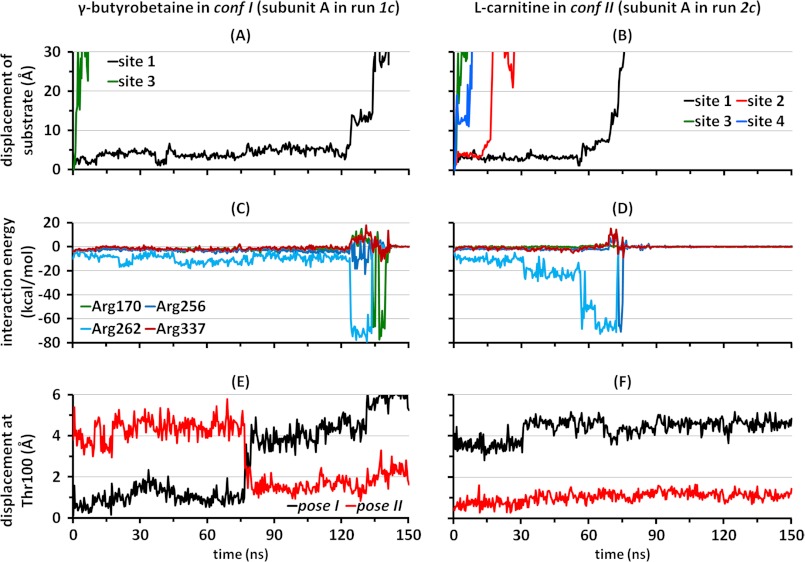
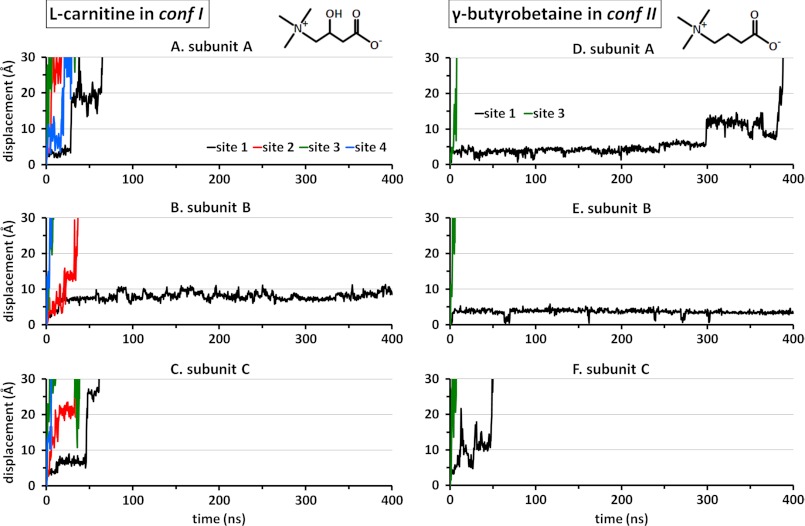
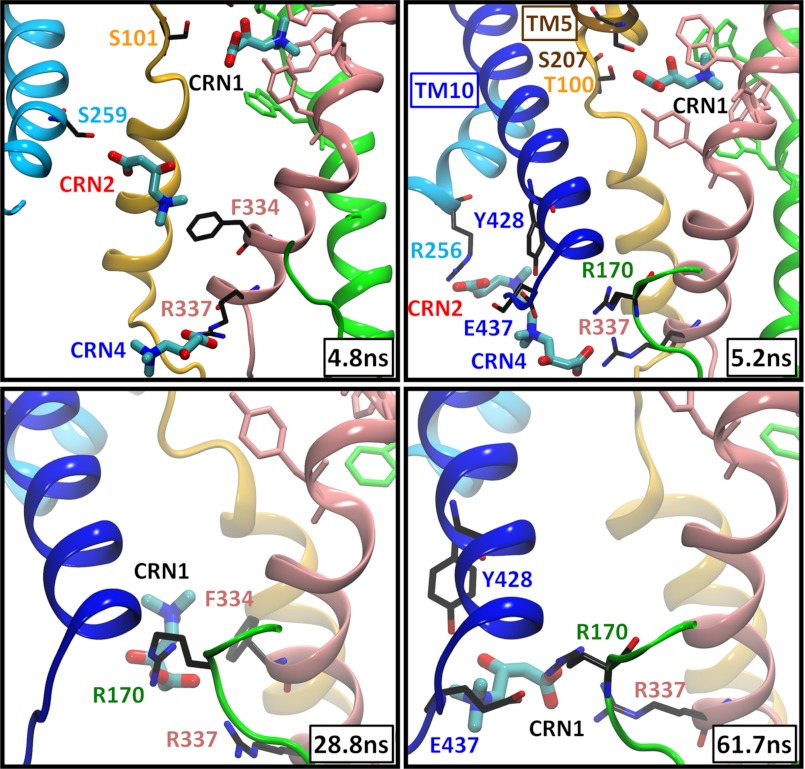
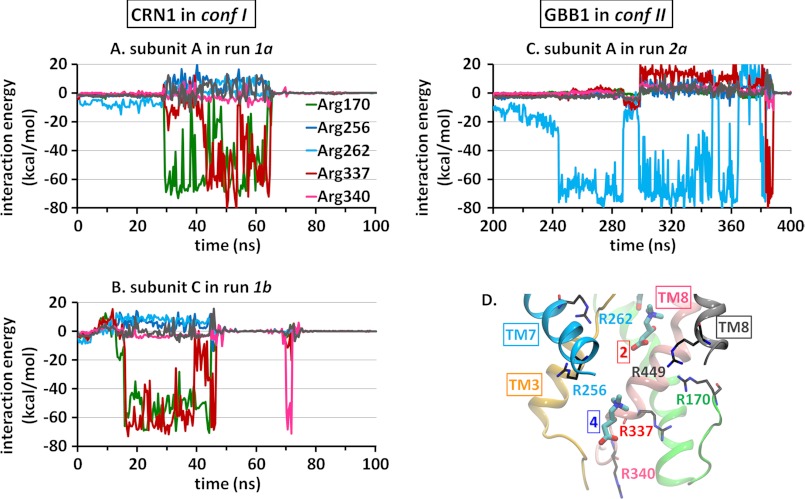
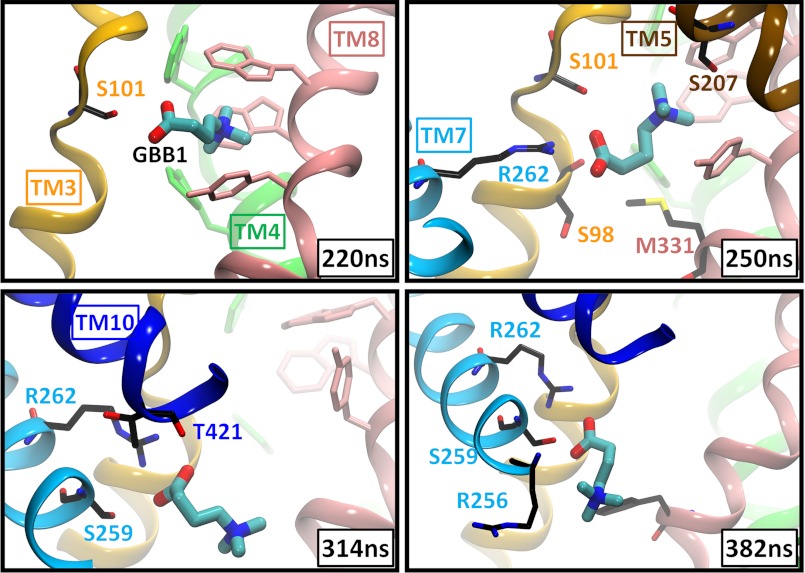
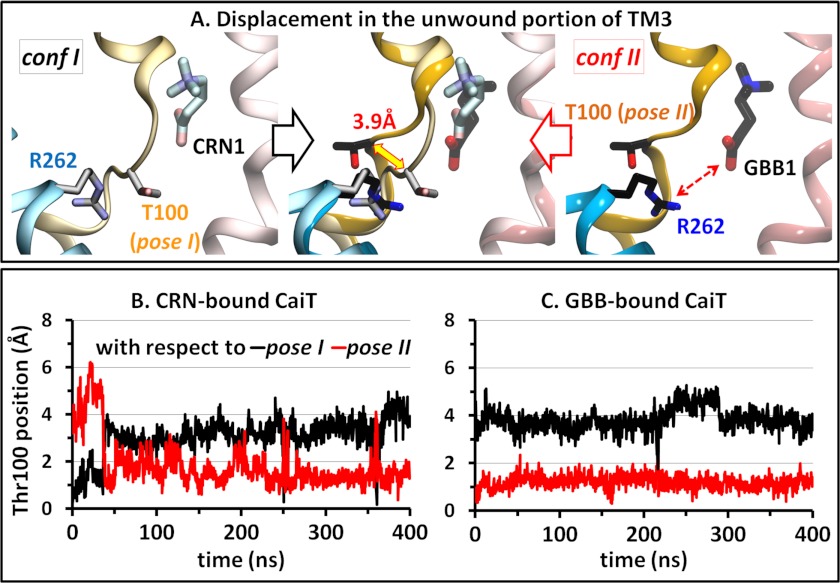
CaiT is a homotrimeric antiporter that exchanges l-carnitine (CRN) with γ-butyrobetaine (GBB) across the bacterial membrane. Three structures have been resolved to date for CaiT, all in the inward-facing state: CRN-bound (with four CRNs per subunit), GBB-bound (two GBBs per subunit), and apo. One of the reported binding sites is the counterpart of the primary site observed in structurally similar transporters. However, the mechanism and pathway(s) of CRN/GBB unbinding and translocation, or even the ability of the substrates to dislodge from the reported binding sites, are yet to be determined. To shed light on these issues, we performed a total of 1.3 μs of molecular dynamics simulations and examined the dynamics of substrate-bound CaiT structures under different conditions. We find that both CRN and GBB are able to dissociate completely from their primary site into the cytoplasm. Substrate molecules initially located at the secondary sites dissociate even faster (within tens of nanoseconds) into the extra- or intracellular regions. Interestingly, the unbinding pathway from the primary site appears to be dictated by the geometry of the unwound part of the transmembrane (TM) helix 3, mostly around Thr(100) therein. Arg(262) on TM7, which apparently mimics the role of Na(+) in CaiT structural homologues, plays a key role in triggering the dissociation of the substrate away from the primary site and guiding its release to the cytoplasm provided that the unwound part of TM3 switches from a shielding to a yielding pose.
CaiT 是一种三聚体反向转运体,可在细菌膜两侧交换左旋肉碱 (CRN) 和γ-丁基甜菜碱 (GBB)。迄今为止,已经解析了 CaiT 的三个结构,均为内向构象:CRN 结合(每个亚基有四个 CRN)、GBB 结合(每个亚基有两个 GBB)和无配体。报道的一个结合位点是在结构相似的转运体中观察到的主要结合位点的对应物。然而,CRN/GBB 结合物的释放和转运机制和途径,甚至是底物从报道的结合位点中脱离的能力,仍有待确定。为了阐明这些问题,我们总共进行了 1.3 μs 的分子动力学模拟,并在不同条件下研究了底物结合的 CaiT 结构的动力学。我们发现 CRN 和 GBB 都能够完全从其主要结合位点解离到细胞质中。最初位于次要结合位点的底物分子甚至更快(在数十纳秒内)解离到细胞外或细胞内区域。有趣的是,从主要结合位点的释放途径似乎由未缠绕跨膜 (TM) 螺旋 3 的部分的几何形状决定,主要是其中的 Thr(100) 附近。TM7 上的 Arg(262) 显然模拟了 CaiT 结构同源物中 Na(+) 的作用,在触发底物从主要结合位点解离并引导其释放到细胞质中起着关键作用,前提是 TM3 的未缠绕部分从屏蔽状态切换到屈服状态。