Graduate School of Agriculture, Tokyo University of Agriculture and Technology (TUAT), Fuchu, Japan.
PLoS One. 2012;7(8):e44101. doi: 10.1371/journal.pone.0044101. Epub 2012 Aug 27.
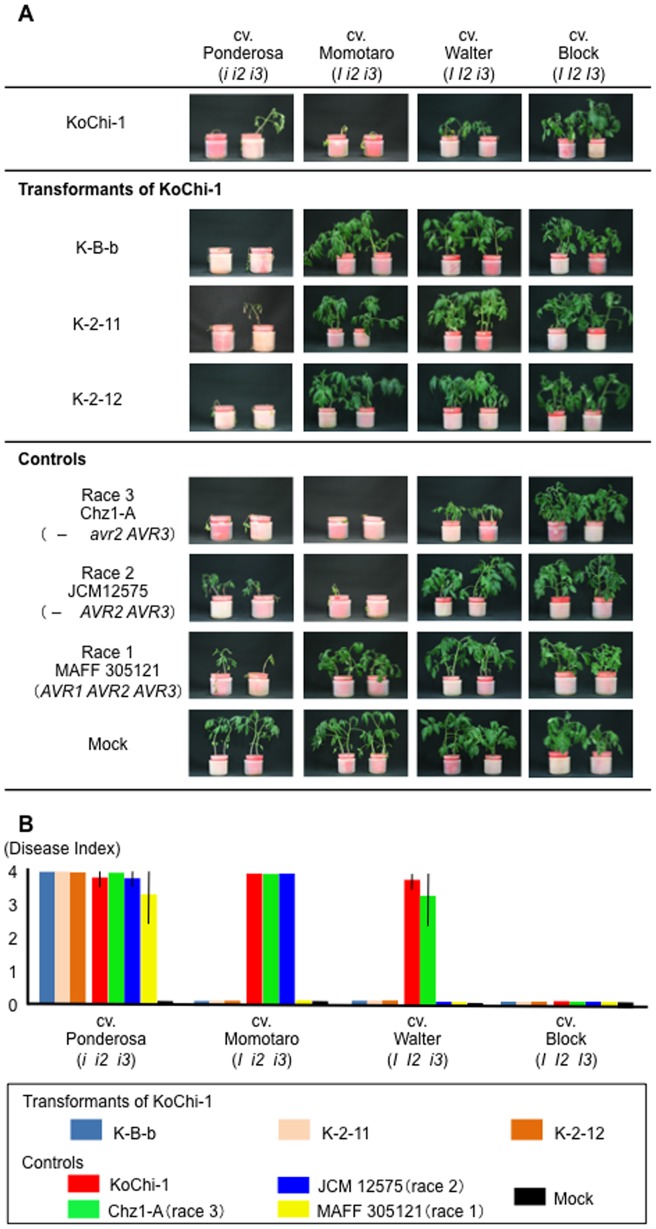
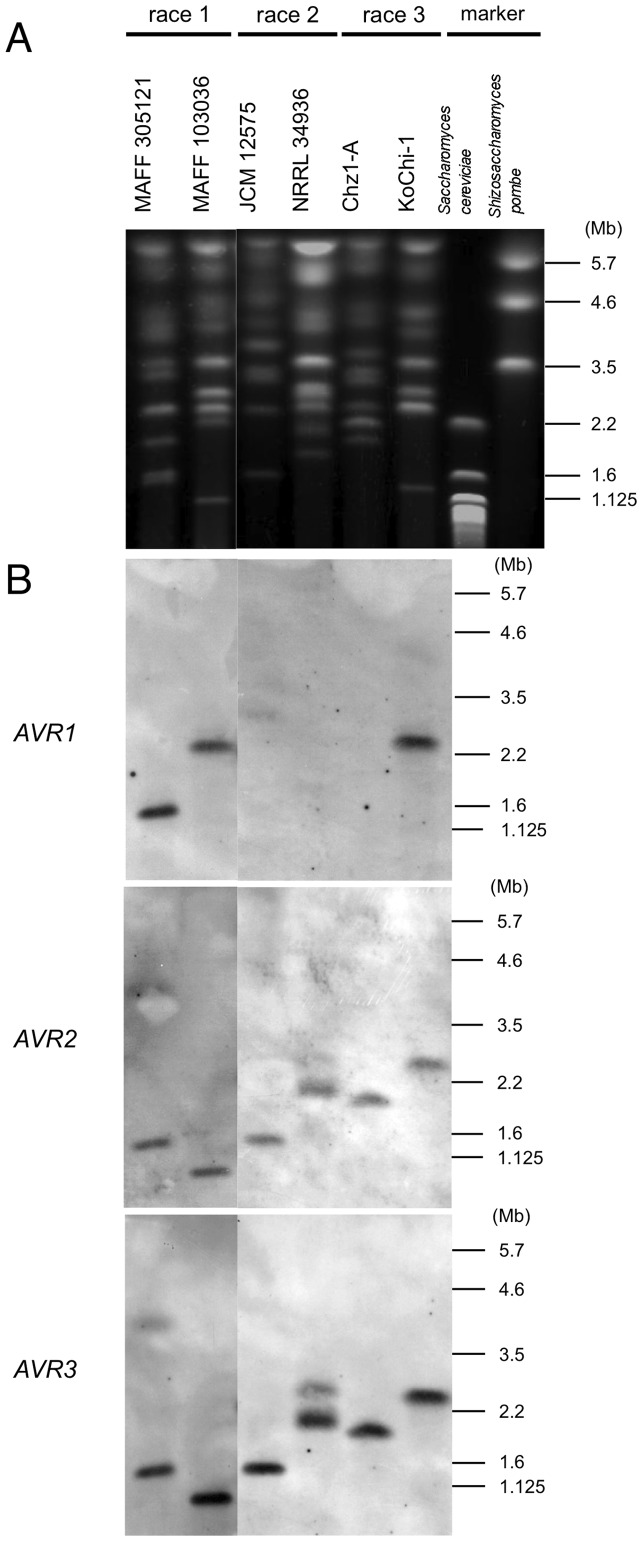
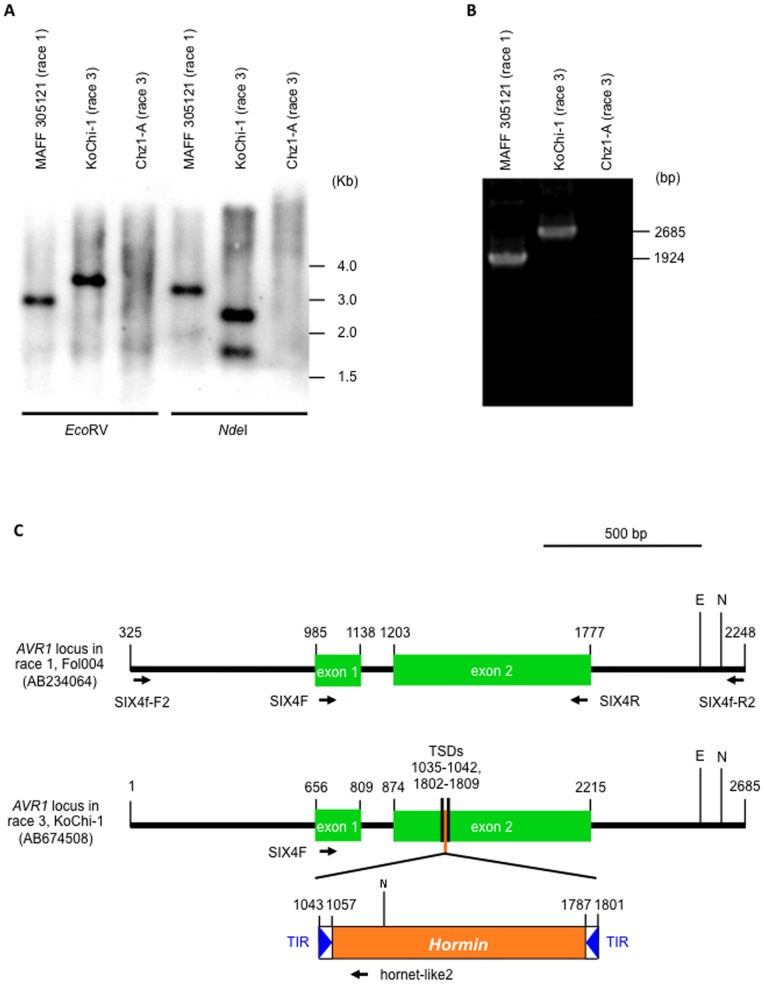
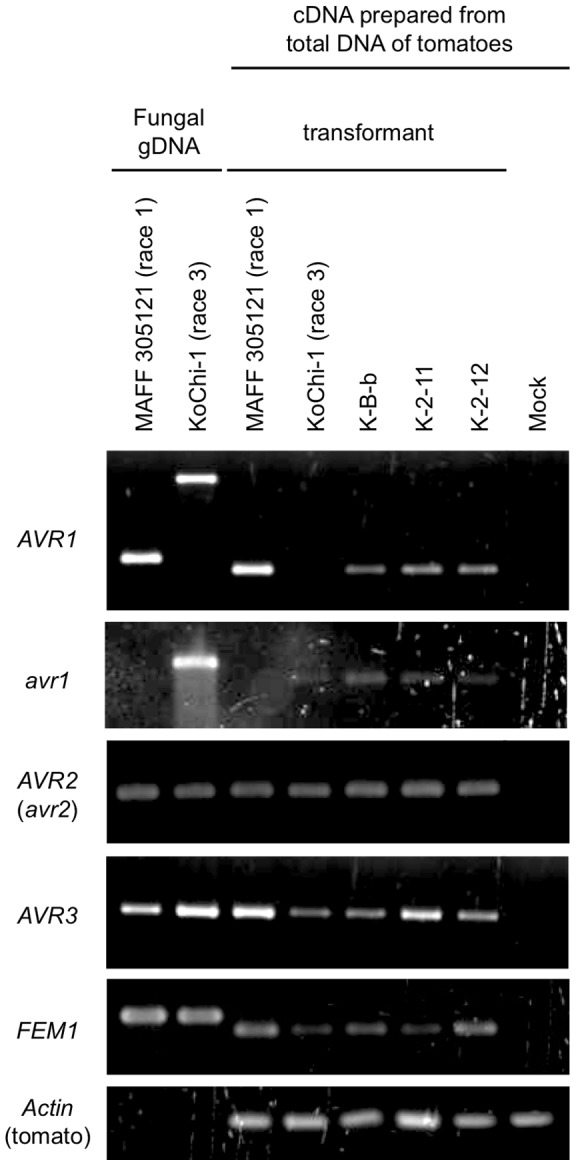
Compatible/incompatible interactions between the tomato wilt fungus Fusarium oxysporum f. sp. lycopersici (FOL) and tomato Solanum lycopersicum are controlled by three avirulence genes (AVR1-3) in FOL and the corresponding resistance genes (I-I3) in tomato. The three known races (1, 2 and 3) of FOL carry AVR genes in different combinations. The current model to explain the proposed order of mutations in AVR genes is: i) FOL race 2 emerged from race 1 by losing the AVR1 and thus avoiding host resistance mediated by I (the resistance gene corresponding to AVR1), and ii) race 3 emerged when race 2 sustained a point mutation in AVR2, allowing it to evade I2-mediated resistance of the host. Here, an alternative mechanism of mutation of AVR genes was determined by analyses of a race 3 isolate, KoChi-1, that we recovered from a Japanese tomato field in 2008. Although KoChi-1 is race 3, it has an AVR1 gene that is truncated by the transposon Hormin, which belongs to the hAT family. This provides evidence that mobile genetic elements may be one of the driving forces underlying race evolution. KoChi-1 transformants carrying a wild type AVR1 gene from race 1 lost pathogenicity to cultivars carrying I, showing that the truncated KoChi-1 avr1 is not functional. These results imply that KoChi-1 is a new race 3 biotype and propose an additional path for emergence of FOL races: Race 2 emerged from race 1 by transposon-insertion into AVR1, not by deletion of the AVR1 locus; then a point mutation in race 2 AVR2 resulted in emergence of race 3.
番茄枯萎病菌(Fusarium oxysporum f. sp. lycopersici,FOL)与番茄(Solanum lycopersicum)之间的亲和/不亲和互作受 FOL 中的三个无毒基因(AVR1-3)和番茄中相应的抗性基因(I-I3)控制。FOL 的已知 3 个小种(1、2 和 3)携带不同组合的 AVR 基因。目前解释 AVR 基因突变顺序的模型为:i)FOL 小种 2 通过失去 AVR1 从而避免由 I 介导的宿主抗性(与 AVR1 对应的抗性基因)而从小种 1 中出现,ii)当小种 2 中的 AVR2 发生点突变时,小种 3 出现,从而使它能够逃避宿主由 I2 介导的抗性。在这里,通过对我们于 2008 年从日本番茄田中回收的小种 3 分离株 KoChi-1 的分析,确定了 AVR 基因突变的另一种机制。尽管 KoChi-1 是小种 3,但它的 AVR1 基因被 Hormin 转座子截断,Hormin 属于 hAT 家族。这表明移动遗传元件可能是小种种群进化的驱动力之一。携带来自小种 1 的野生型 AVR1 基因的 KoChi-1 转化体对携带 I 的品种失去致病性,表明截断的 KoChi-1 avr1 没有功能。这些结果表明 KoChi-1 是一种新的小种 3 生物型,并提出了 FOL 小种出现的另一种途径:小种 2 从小种 1 通过 AVR1 插入转座子而不是 AVR1 基因座缺失而出现;然后小种 2 的 AVR2 发生点突变导致小种 3 出现。