Institute of Molecular Life Sciences, University of Zurich, Winterthurerstrasse 190, Zurich, CH 8057, Switzerland.
Neural Dev. 2012 Nov 9;7:36. doi: 10.1186/1749-8104-7-36.
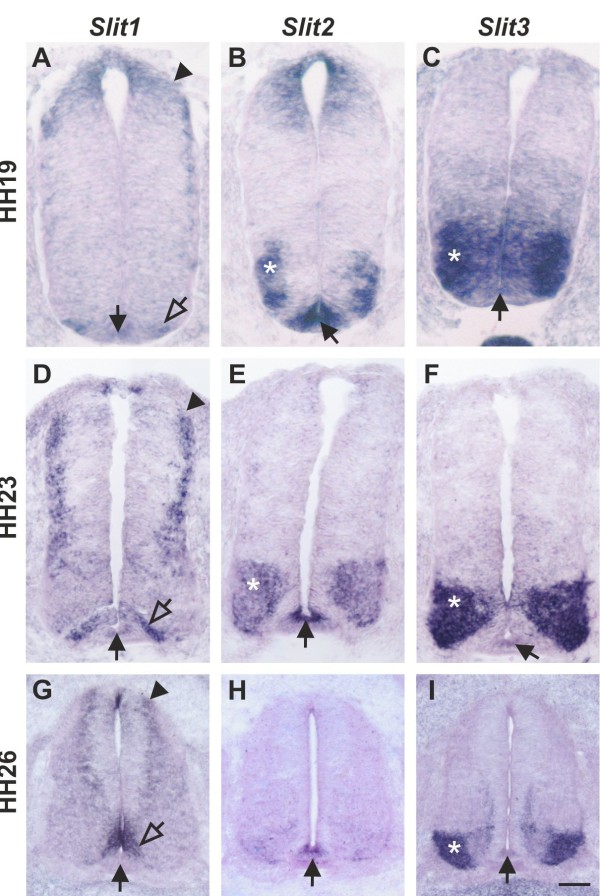
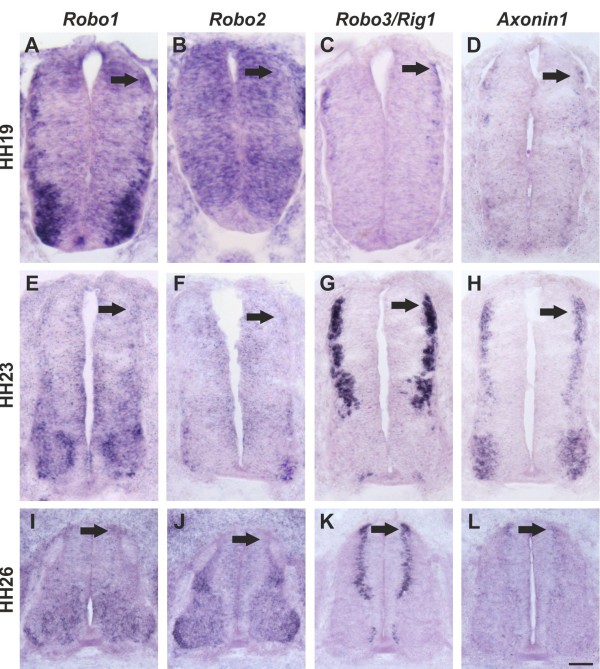
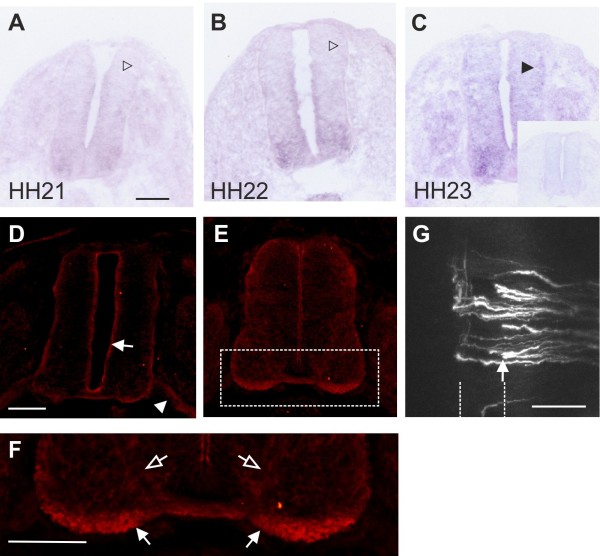
Axons navigate to their future synaptic targets with the help of choice points, intermediate targets that express axon guidance cues. Once they reach a choice point, axons need to switch their response from attraction to repulsion in order to move on with the next stage of their journey. The mechanisms underlying the change in axonal responsiveness are poorly understood. Commissural axons become sensitive to the repulsive activity of Slits when they cross the ventral midline of the CNS. Responsiveness to Slits depends on surface expression of Robo receptors. In Drosophila, Commissureless (Comm) plays a crucial regulatory role in midline crossing by keeping Robo levels low on precommissural axons. Interestingly, to date no vertebrate homolog of comm has been identified. Robo3/Rig1 has been shown to control Slit sensitivity before the midline, but without affecting Robo1 surface expression.
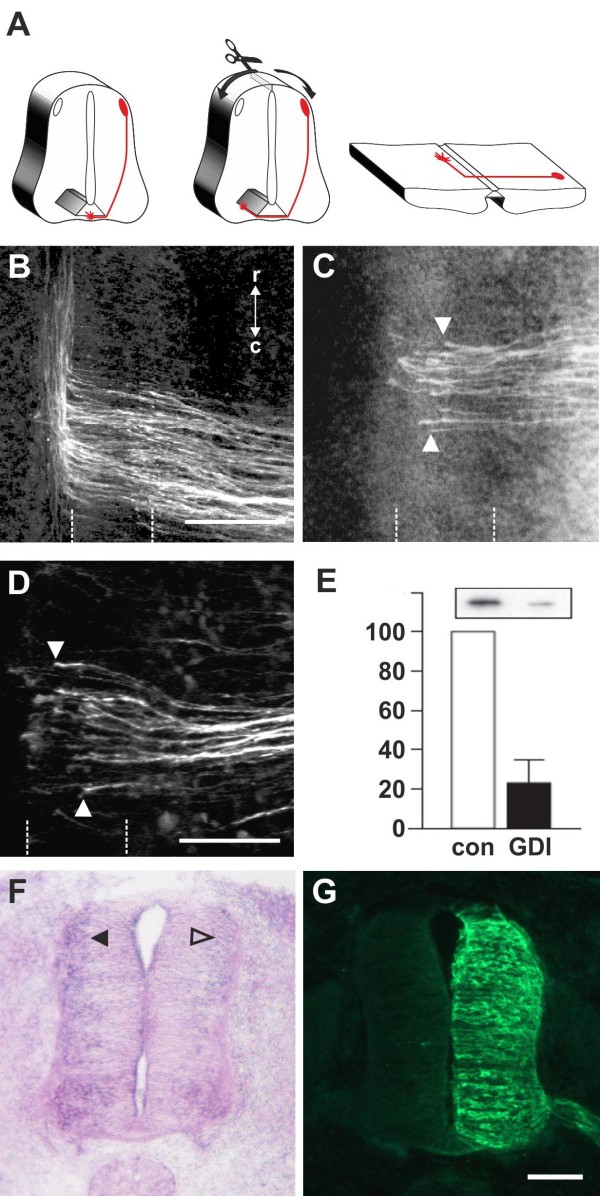
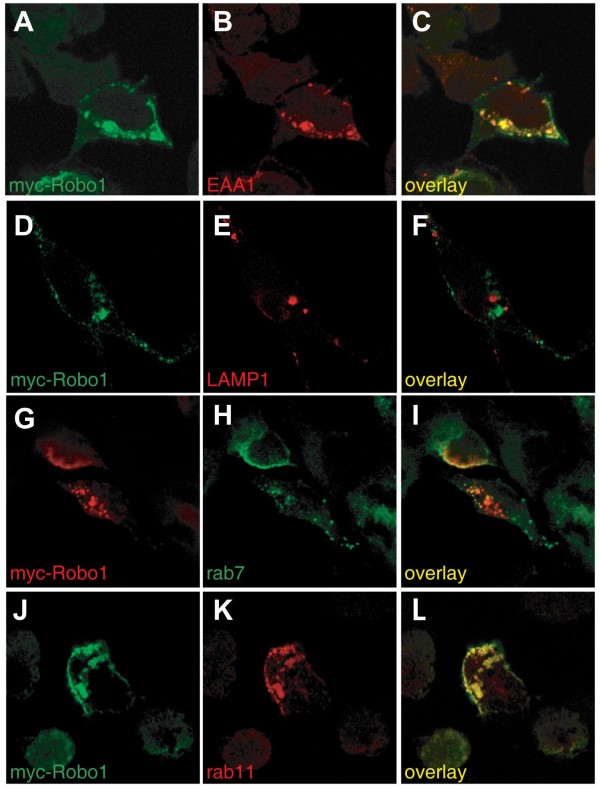
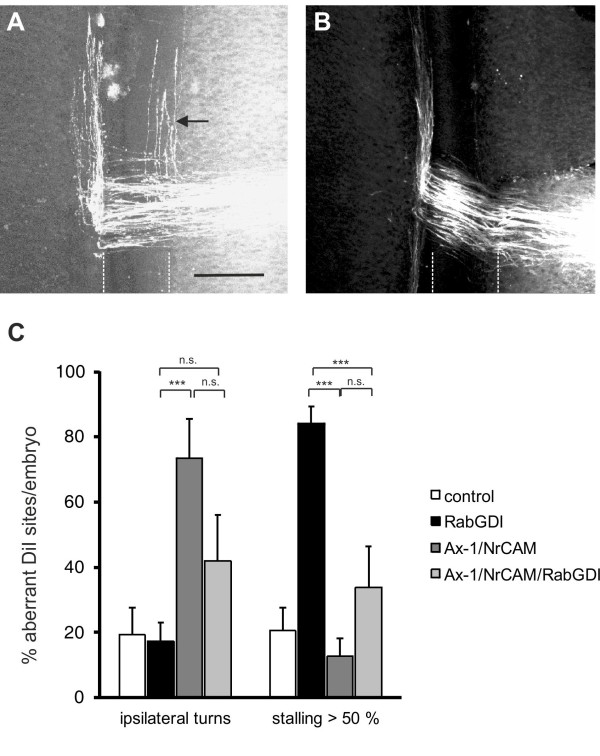
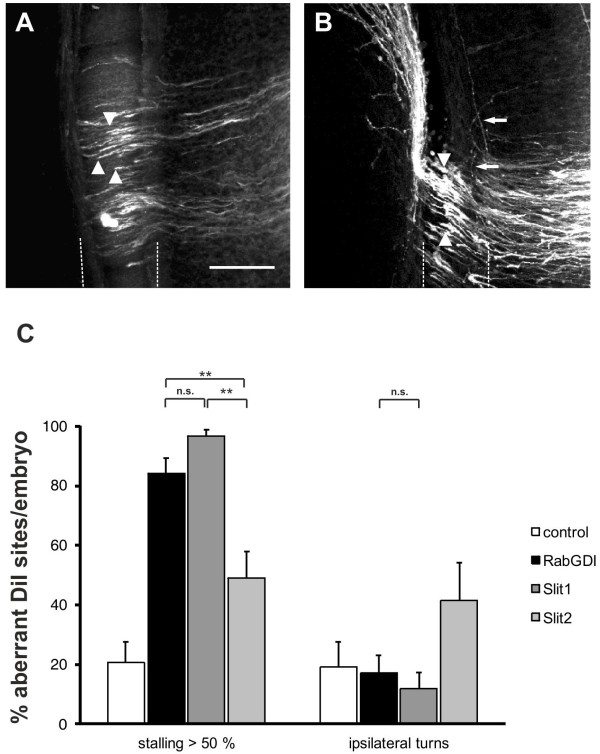
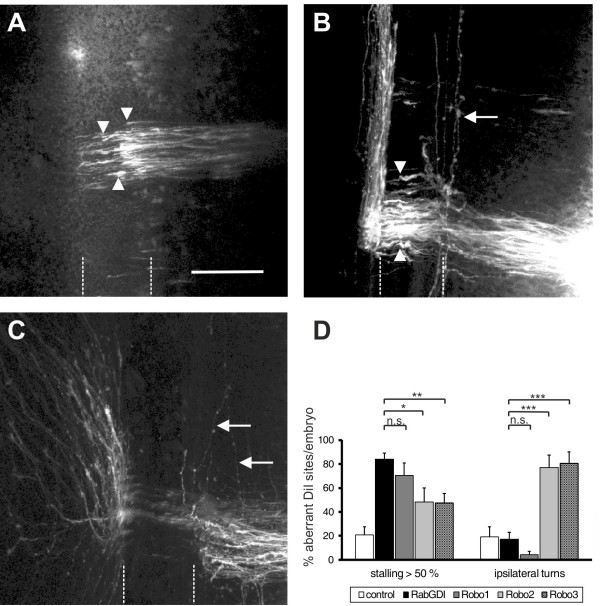
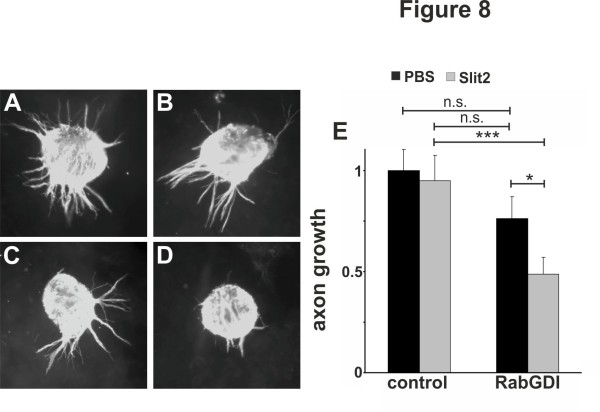
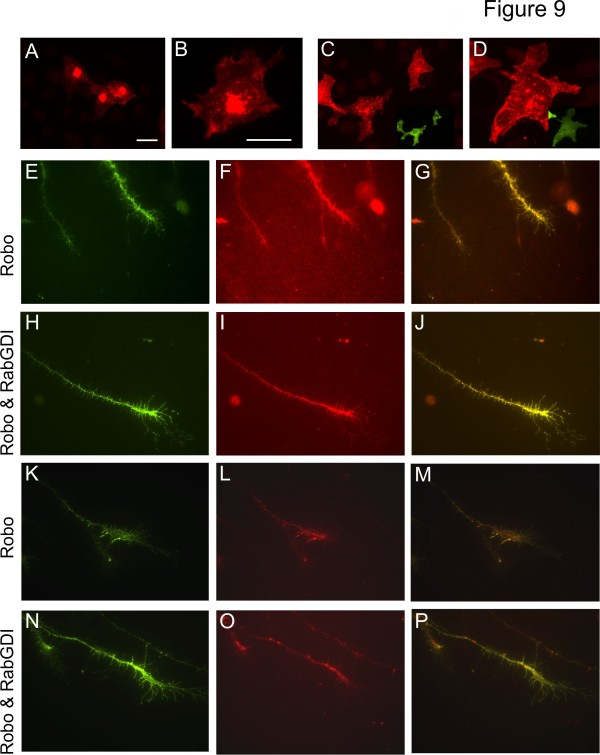
We had identified RabGDI, a gene linked to human mental retardation and an essential component of the vesicle fusion machinery, in a screen for differentially expressed floor-plate genes. Downregulation of RabGDI by in ovo RNAi caused commissural axons to stall in the floor plate, phenocopying the effect observed after downregulation of Robo1. Conversely, premature expression of RabGDI prevented commissural axons from entering the floor plate. Furthermore, RabGDI triggered Robo1 surface expression in cultured commissural neurons. Taken together, our results identify RabGDI as a component of the switching mechanism that is required for commissural axons to change their response from attraction to repulsion at the intermediate target.
RabGDI takes over the functional role of fly Comm by regulating the surface expression of Robo1 on commissural axons in vertebrates. This in turn allows commissural axons to switch from attraction to repulsion at the midline of the spinal cord.
轴突在选择点的帮助下向未来的突触靶标导航,选择点是表达轴突导向线索的中间靶标。一旦它们到达选择点,轴突就需要将其反应从吸引转变为排斥,以便继续进行旅程的下一个阶段。轴突反应性变化的机制还知之甚少。当 Commissural 轴突穿过 CNS 的腹中线时,它们对 Slits 的排斥活性变得敏感。对 Slits 的反应性取决于 Robo 受体的表面表达。在果蝇中,Commissureless (Comm) 通过保持 precommissural 轴突上 Robo 水平低,在中线穿越中发挥关键的调节作用。有趣的是,到目前为止,还没有鉴定出 Comm 的脊椎动物同源物。Robo3/Rig1 已被证明在中线之前控制 Slit 敏感性,但不影响 Robo1 表面表达。
我们在筛选差异表达的基板基因时发现了 RabGDI,这是一个与人类智力迟钝相关的基因,也是囊泡融合机制的重要组成部分。在鸡胚中的 RNAi 下调 RabGDI 会导致 Commissural 轴突在基板中停滞,这与 Robo1 下调后观察到的效果相似。相反,RabGDI 的过早表达会阻止 Commissural 轴突进入基板。此外,RabGDI 在培养的 Commissural 神经元中触发 Robo1 的表面表达。总之,我们的结果表明 RabGDI 是切换机制的一个组成部分,该机制要求 Commissural 轴突在中间靶标处将其反应从吸引转变为排斥。
RabGDI 通过调节脊椎动物 Commissural 轴突上 Robo1 的表面表达,取代了果蝇 Comm 的功能作用。这反过来又允许 Commissural 轴突在脊髓中线从吸引转变为排斥。