Department of Genetics, University of Cambridge, Downing Site, Downing Street, Cambridge CB2 3EH, UK.
Open Biol. 2012 Nov;2(11):120132. doi: 10.1098/rsob.120132.
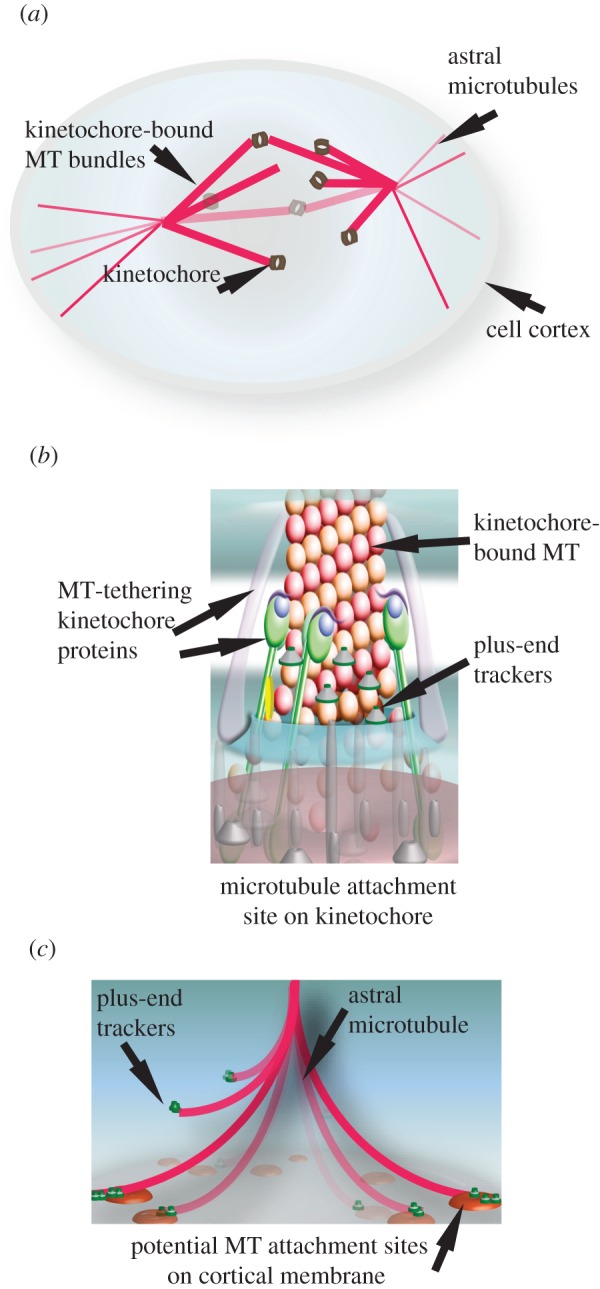
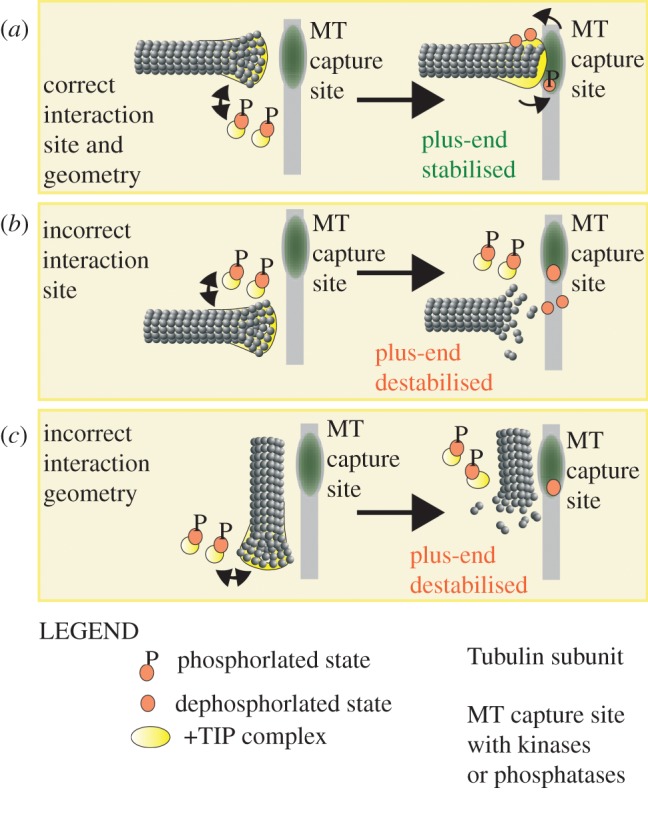
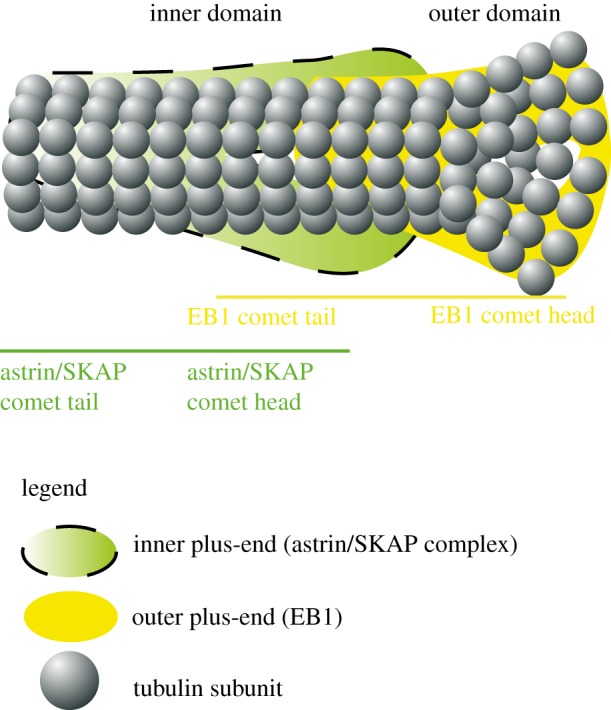
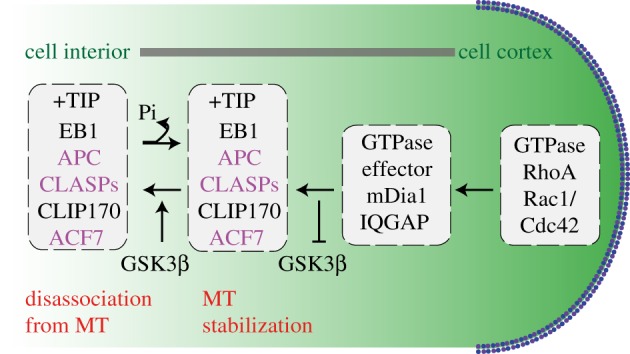
The microtubule polymer grows and shrinks predominantly from one of its ends called the 'plus-end'. Plus-end regulation during interphase is well understood. However, mitotic regulation of plus-ends is only beginning to be understood in mammalian cells. During mitosis, the plus-ends are tethered to specialized microtubule capture sites. At these sites, plus-end-binding proteins are loaded and unloaded in a regulated fashion. Proper tethering of plus-ends to specialized sites is important so that the microtubule is able to translate its growth and shrinkage into pushing and pulling forces that move bulky subcellular structures. We discuss recent advances on how mitotic plus-ends are tethered to distinct subcellular sites and how plus-end-bound proteins can modulate the forces that move subcellular structures. Using end binding 1 (EB1) as a prototype plus-end-binding protein, we highlight the complex network of plus-end-binding proteins and their regulation through phosphorylation. Finally, we develop a speculative 'moving platform' model that illustrates the plus-end's role in distinguishing correct versus incorrect microtubule interactions.
微管聚合物主要从其称为“正端”的一个末端生长和收缩。 间期的正端调控已经得到很好的理解。 然而,有丝分裂过程中对正端的调控才刚刚开始在哺乳动物细胞中被理解。 在有丝分裂过程中,正端被固定在专门的微管捕获位点上。 在这些位点上,正端结合蛋白以受调控的方式加载和卸载。 正端与专门的位点适当固定非常重要,这样微管才能将其生长和收缩转化为推动和拉动的力,从而移动体积较大的亚细胞结构。 我们讨论了有丝分裂正端与不同亚细胞位点固定的最新进展,以及正端结合蛋白如何调节移动亚细胞结构的力。 我们使用末端结合蛋白 1(EB1)作为正端结合蛋白的原型,强调了正端结合蛋白及其通过磷酸化调节的复杂网络。 最后,我们提出了一个推测性的“移动平台”模型,说明了正端在区分正确和错误的微管相互作用中的作用。