The Division of Immunology, Department of Medicine, Weill Medical College, Cornell University New York, NY, USA.
Front Immunol. 2012 Nov 29;3:364. doi: 10.3389/fimmu.2012.00364. eCollection 2012.
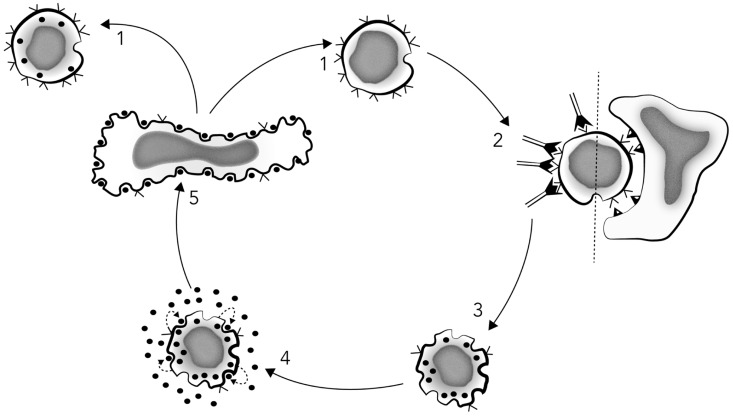
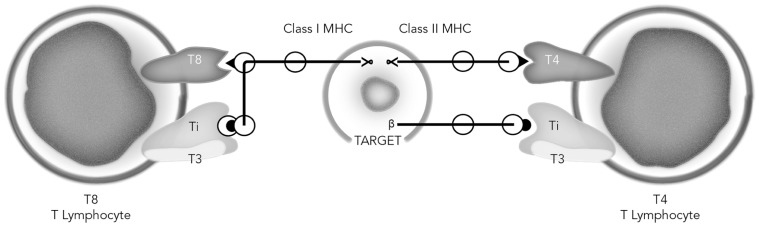
By 1980 it was obvious that to more fully understand adaptive immunity, one needed to somehow reduce the tremendous complexity of antigen recognition by T cell populations. Thus, there were two developments that resulted in a paradigm shift in immunology, one being the generation of monoclonal antibodies (MoAbs), and the other the development of monoclonal functional antigen-specific T cell lines. For the first time, the cellular reagents became available to ask new questions as to how individual cells comprising the complex cell populations recognize and respond to changes in their molecular environments. The first successful generation of monoclonal T cells depended upon the understanding that antigen renders cells responsive to the antigen non-specific T cell growth factor that came to be termed interleukin-2 (IL-2), which could then be used in propagating large numbers of the progeny of single cells, which in turn could then be used for molecular analyses. Monoclonal functional human T cells were used to immunize mice to generate clone-specific (clonotypic) MoAbs, which then permitted the first biochemical characterizations of the antigen recognition elements of the T cell antigen receptor (TCR) complex. Moreover, the use of monoclonal cytolytic and helper/inducer human T cell clones essentially proved that the T cell-specific molecules T4 (CD4) and T8 (CD8) functioned as accessory molecules in antigen recognition by defining MHC class II or class I restriction respectively. As well, the expression of the T3 (CD3) molecules, found to be common to all T cells, were shown further to be obligatory for functional antigen-specific T cell signaling. The monoclonal IL-2-dependent T cells were also instrumental in the isolation and purification of the IL-2 molecule to homogeneity, the first interleukin molecule to be identified and characterized. These advances then led to the generation of pure radiolabeled IL-2 molecules that were used to identify the first interleukin cellular receptors, and as well the generation of the first MoAbs reactive with both IL-2 and IL-2 receptors. All of these advances led subsequently to the isolation of the first cDNA clones recognizing one of the two chains comprising the T cell antigen recognition elements (β-chain), as well cDNA clones encoding IL-2. Accordingly, armed with all of these unique cellular and molecular reagents, it was possible to determine that antigen triggering of the TCR complex initiates IL-2 production and IL-2 receptor expression, which in turn initiate the T cell clonal proliferative expansion, envisioned by Burnet in his formulation of the clonal selection theory. Thus, adaptive immunity receives antigen-specific activation signals from the environment and turns them into antigen non-specific endogenous action signals.
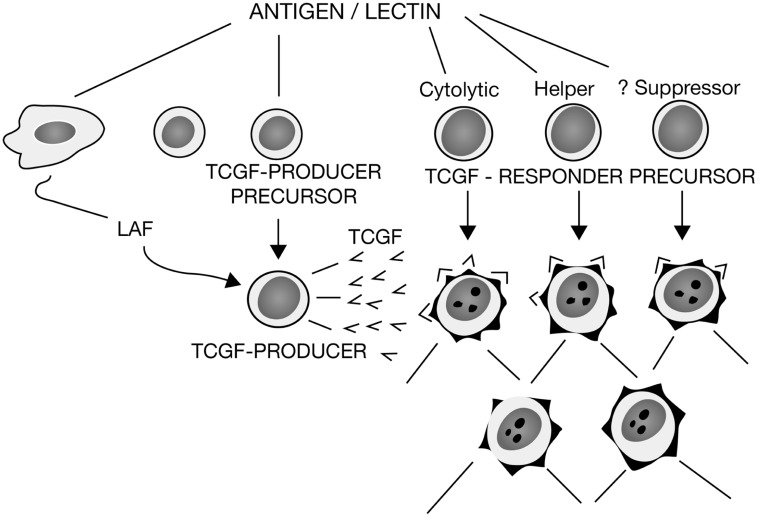
到 1980 年,人们已经清楚地认识到,如果要更全面地了解适应性免疫,就需要以某种方式降低 T 细胞群体对抗原识别的巨大复杂性。因此,有两个发展导致了免疫学的范式转变,一个是单克隆抗体 (MoAb) 的产生,另一个是单克隆功能性抗原特异性 T 细胞系的发展。首次获得了细胞试剂,可以提出新的问题,即组成复杂细胞群体的单个细胞如何识别和响应其分子环境的变化。第一代成功的单克隆 T 细胞的产生依赖于这样的理解,即抗原使细胞对非特异性 T 细胞生长因子(后来被称为白细胞介素 2 (IL-2))产生反应,然后可以使用这种生长因子来繁殖大量单细胞的后代,这些后代又可以用于分子分析。单克隆功能性人 T 细胞被用于免疫小鼠以产生克隆特异性(克隆型)MoAb,然后可以对 T 细胞抗原受体 (TCR) 复合物的抗原识别元件进行首次生化表征。此外,使用单克隆细胞毒性和辅助/诱导人 T 细胞克隆实质上证明了 T 细胞特异性分子 T4(CD4)和 T8(CD8)分别作为辅助分子在抗原识别中起作用,分别定义了 MHC Ⅱ类或Ⅰ类限制。同样,发现存在于所有 T 细胞中的 T3(CD3)分子的表达进一步表明对于功能性抗原特异性 T 细胞信号传导是必需的。单克隆 IL-2 依赖性 T 细胞对于 IL-2 分子的分离和纯化也很重要,这是第一个被鉴定和表征的白细胞介素分子。这些进展随后导致了纯放射性标记的 IL-2 分子的产生,这些分子用于鉴定第一个白细胞介素细胞受体,并产生与 IL-2 和 IL-2 受体都反应的第一个 MoAb。所有这些进展随后导致了第一个识别 TCR 复合物的两个链之一的 cDNA 克隆的分离(β链),以及编码 IL-2 的 cDNA 克隆。因此,有了所有这些独特的细胞和分子试剂,就有可能确定抗原触发 TCR 复合物会引发 IL-2 的产生和 IL-2 受体的表达,而这反过来又会引发 Burnet 在其克隆选择理论中所设想的 T 细胞克隆增殖扩张。因此,适应性免疫从环境中接收到抗原特异性激活信号,并将其转化为抗原非特异性内源性作用信号。