Department of Pharmaceutical Chemistry & Bioanalytics, Institute of Pharmacy, Martin-Luther University Halle-Wittenberg, Halle, Saale, Germany.
PLoS One. 2013 Apr 25;8(4):e61913. doi: 10.1371/journal.pone.0061913. Print 2013.
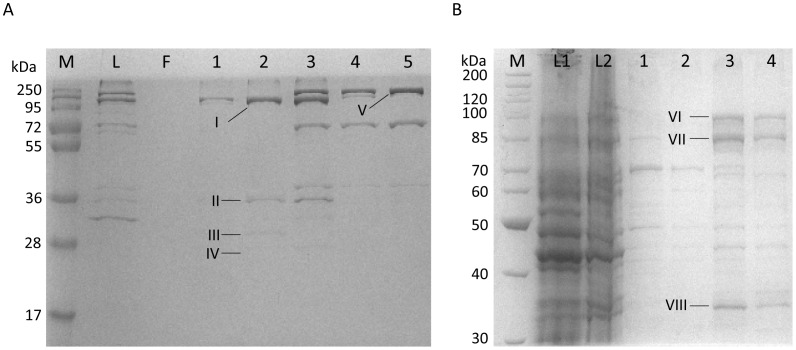
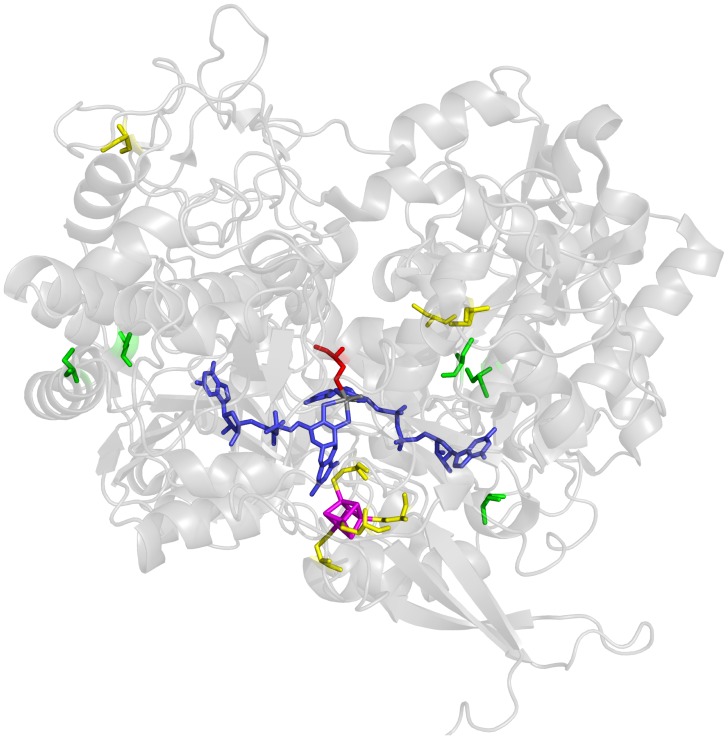
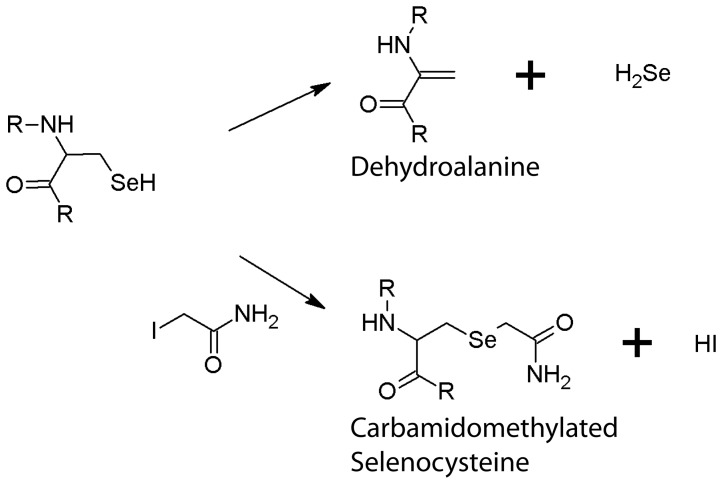
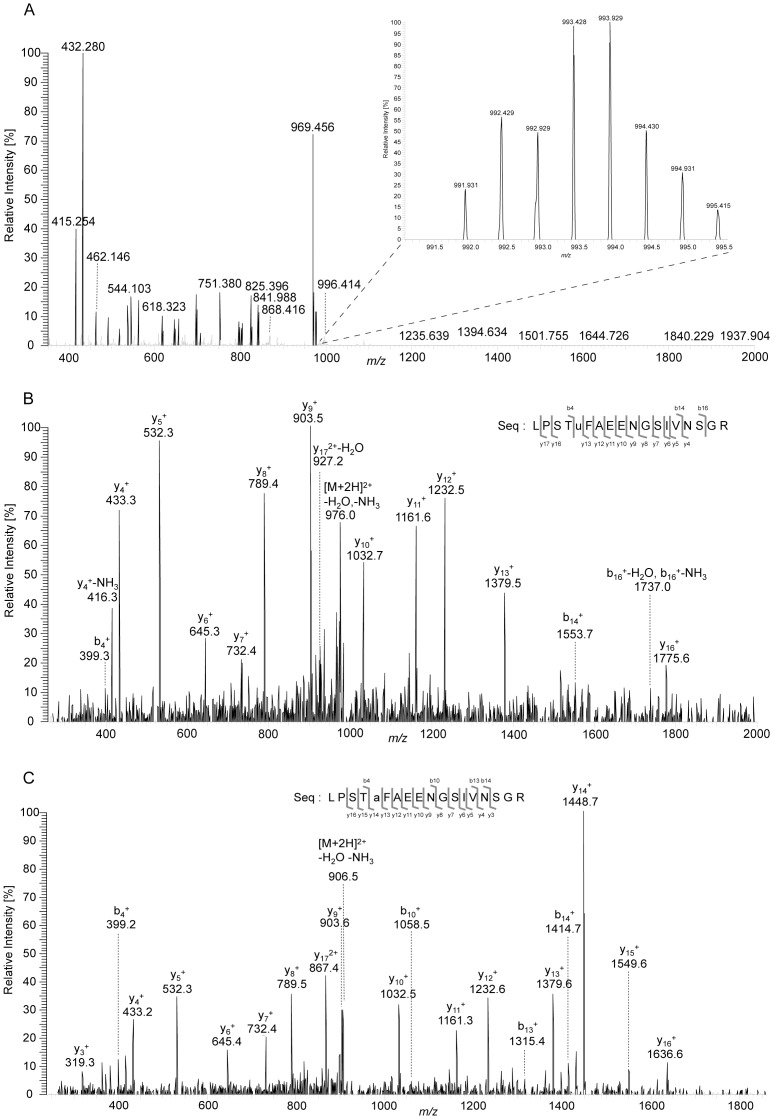
The formate dehydrogenases (Fdh) Fdh-O, Fdh-N, and Fdh-H, are the only proteins in Escherichia coli that incorporate selenocysteine at a specific position by decoding a UGA codon. However, an excess of selenium can lead to toxicity through misincorporation of selenocysteine into proteins. To determine whether selenocysteine substitutes for cysteine, we grew Escherichia coli in the presence of excess sodium selenite. The respiratory Fdh-N and Fdh-O enzymes, along with nitrate reductase (Nar) were co-purified from wild type strain MC4100 after anaerobic growth with nitrate and either 2 µM or 100 µM selenite. Mass spectrometric analysis of the catalytic subunits of both Fdhs identified the UGA-specified selenocysteine residue and revealed incorporation of additional, 'non-specific' selenocysteinyl residues, which always replaced particular cysteinyl residues. Although variable, their incorporation was not random and was independent of the selenite concentration used. Notably, these cysteines are likely to be non-essential for catalysis and they do not coordinate the iron-sulfur cluster. The remaining cysteinyl residues that could be identified were never substituted by selenocysteine. Selenomethionine was never observed in our analyses. Non-random substitution of particular cysteinyl residues was also noted in the electron-transferring subunit of both Fdhs as well as in the subunits of the Nar enzyme. Nar isolated from an E. coli selC mutant also showed a similar selenocysteine incorporation pattern to the wild-type indicating that non-specific selenocysteine incorporation was independent of the specific selenocysteine pathway. Thus, selenide replaces sulfide in the biosynthesis of cysteine and misacylated selenocysteyl-tRNA(Cys) decodes either UGU or UGC codons, which usually specify cysteine. Nevertheless, not every UGU or UGC codon was decoded as selenocysteine. Together, our results suggest that a degree of misincorporation of selenocysteine into enzymes through replacement of particular, non-essential cysteines, is tolerated and this might act as a buffering system to cope with excessive intracellular selenium.
甲酸脱氢酶(Fdh)Fdh-O、Fdh-N 和 Fdh-H 是大肠杆菌中唯一通过解码 UGA 密码子将硒代半胱氨酸掺入特定位置的蛋白质。然而,过量的硒会导致毒性,因为硒代半胱氨酸会错误地掺入蛋白质中。为了确定硒代半胱氨酸是否可以替代半胱氨酸,我们在过量亚硒酸钠存在的情况下培养大肠杆菌。从厌氧生长硝酸盐并用 2 µM 或 100 µM 亚硒酸钠共纯化野生型菌株 MC4100 的呼吸 Fdh-N 和 Fdh-O 酶以及硝酸还原酶(Nar)后,对两种 Fdh 的催化亚基进行了质谱分析,鉴定了 UGA 指定的硒代半胱氨酸残基,并揭示了额外的“非特异性”硒代半胱氨酸残基的掺入,这些残基总是取代特定的半胱氨酸残基。尽管变化不定,但它们的掺入不是随机的,也与使用的亚硒酸钠浓度无关。值得注意的是,这些半胱氨酸可能对催化作用不是必需的,并且它们不与铁硫簇配位。可以识别的其余半胱氨酸残基从未被硒代半胱氨酸取代。在我们的分析中从未观察到硒甲硫氨酸。在两种 Fdh 的电子传递亚基以及 Nar 酶的亚基中也注意到特定半胱氨酸残基的非随机取代。从大肠杆菌 selC 突变体中分离的 Nar 也表现出与野生型相似的硒代半胱氨酸掺入模式,表明非特异性硒代半胱氨酸掺入独立于特定的硒代半胱氨酸途径。因此,硒化物在半胱氨酸的生物合成中取代硫,错误酰化的硒代半胱氨酸-tRNA(Cys)解码 UGU 或 UGC 密码子,这些密码子通常指定半胱氨酸。然而,并非每个 UGU 或 UGC 密码子都被解码为硒代半胱氨酸。总之,我们的结果表明,通过替换特定的非必需半胱氨酸,将一定程度的硒代半胱氨酸错误掺入酶中是可以容忍的,这可能是一种缓冲系统,可以应对细胞内过量的硒。