Department of Plant Pathology, University of Wisconsin-Madison, Madison, Wisconsin, United States of America.
PLoS Pathog. 2013;9(4):e1003313. doi: 10.1371/journal.ppat.1003313. Epub 2013 Apr 18.
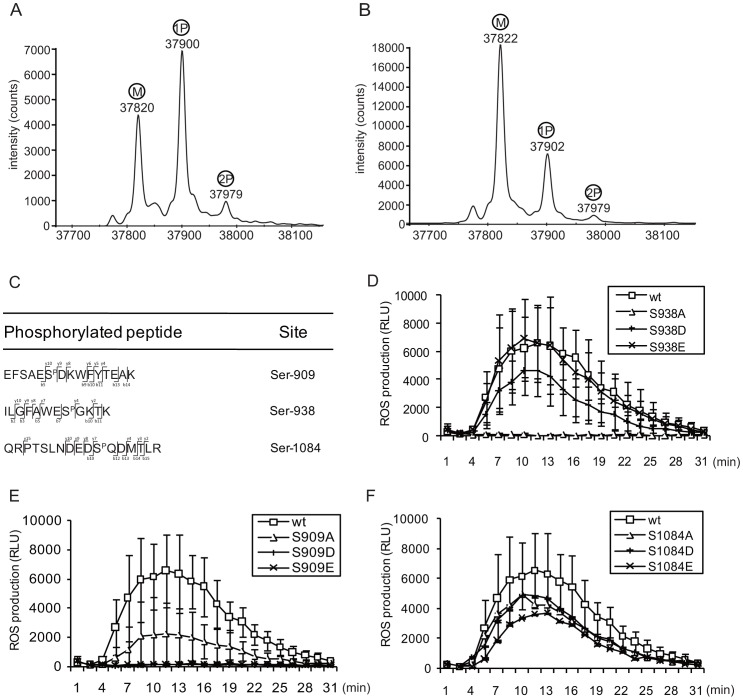
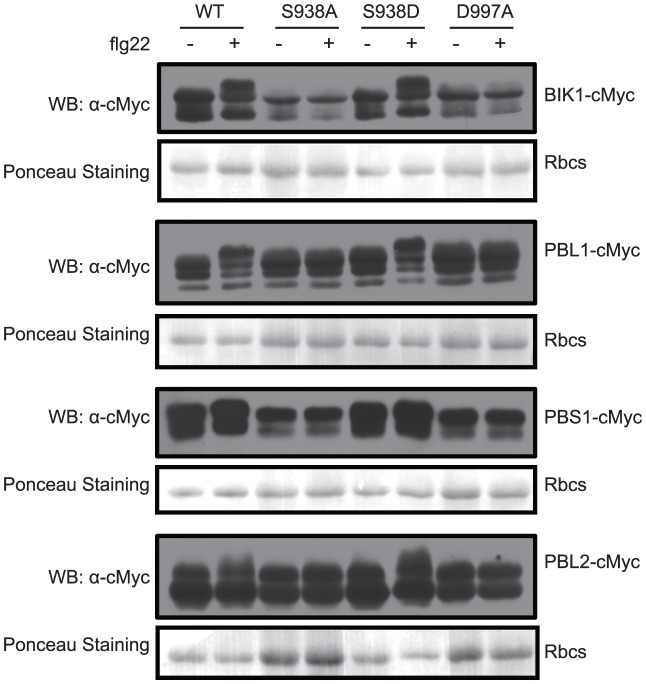
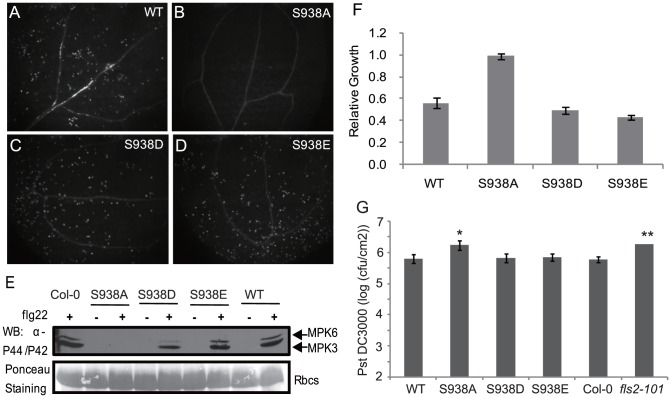
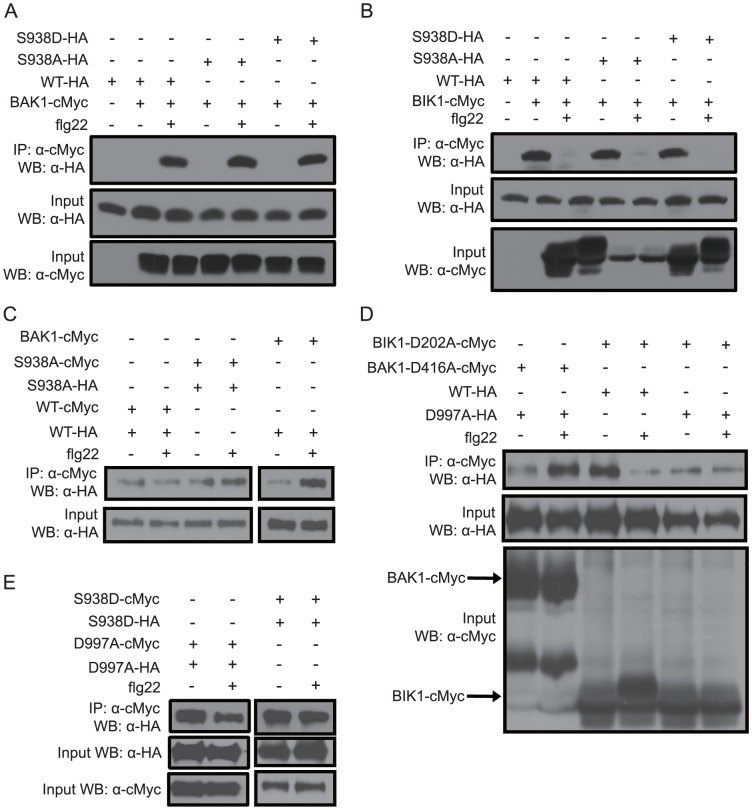
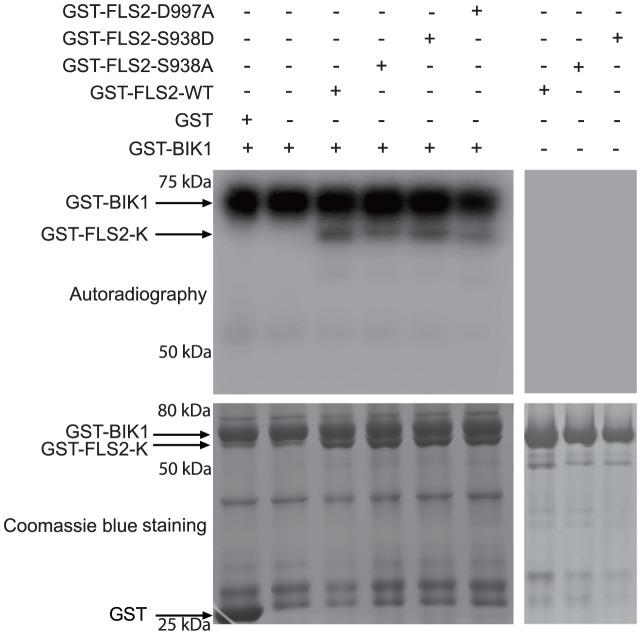
Flagellin-sensing 2 (FLS2) is a leucine-rich repeat/transmembrane domain/protein kinase (LRR-RLK) that is the plant receptor for bacterial flagellin or the flagellin-derived flg22 peptide. Previous work has shown that after flg22 binding, FLS2 releases BIK1 kinase and homologs and associates with BAK1 kinase, and that FLS2 kinase activity is critical for FLS2 function. However, the detailed mechanisms for activation of FLS2 signaling remain unclear. The present study initially identified multiple FLS2 in vitro phosphorylation sites and found that Serine-938 is important for FLS2 function in vivo. FLS2-mediated immune responses are abolished in transgenic plants expressing FLS2(S938A), while the acidic phosphomimic mutants FLS2(S938D) and FLS2(S938E) conferred responses similar to wild-type FLS2. FLS2-BAK1 association and FLS2-BIK1 disassociation after flg22 exposure still occur with FLS2(S938A), demonstrating that flg22-induced BIK1 release and BAK1 binding are not sufficient for FLS2 activity, and that Ser-938 controls other aspects of FLS2 activity. Purified BIK1 still phosphorylated purified FLS2(S938A) and FLS2(S938D) mutant kinase domains in vitro. Phosphorylation of BIK1 and homologs after flg22 exposure was disrupted in transgenic Arabidopsis thaliana plants expressing FLS2(S938A) or FLS2(D997A) (a kinase catalytic site mutant), but was normally induced in FLS2(S938D) plants. BIK1 association with FLS2 required a kinase-active FLS2, but FLS2-BAK1 association did not. Hence FLS2-BIK1 dissociation and FLS2-BAK1 association are not sufficient for FLS2-mediated defense activation, but the proposed FLS2 phosphorylation site Ser-938 and FLS2 kinase activity are needed both for overall defense activation and for appropriate flg22-stimulated phosphorylation of BIK1 and homologs.
Flagellin-sensing 2 (FLS2) 是一种富含亮氨酸重复序列/跨膜结构域/蛋白激酶(LRR-RLK),是植物对细菌鞭毛蛋白或鞭毛蛋白衍生的 flg22 肽的受体。先前的研究表明,在 flg22 结合后,FLS2 释放 BIK1 激酶及其同源物,并与 BAK1 激酶结合,并且 FLS2 激酶活性对于 FLS2 功能至关重要。然而,FLS2 信号转导的激活的详细机制仍不清楚。本研究最初鉴定了多个 FLS2 的体外磷酸化位点,并发现丝氨酸-938对于 FLS2 在体内的功能很重要。在表达 FLS2(S938A)的转基因植物中,FLS2 介导的免疫反应被消除,而酸性磷酸模拟突变体 FLS2(S938D)和 FLS2(S938E)赋予与野生型 FLS2 相似的反应。在 flg22 暴露后,FLS2-BAK1 结合和 FLS2-BIK1 解离仍然发生在 FLS2(S938A)中,表明 flg22 诱导的 BIK1 释放和 BAK1 结合不足以激活 FLS2,而 Ser-938 控制 FLS2 活性的其他方面。纯化的 BIK1 仍能在体外磷酸化纯化的 FLS2(S938A)和 FLS2(S938D)突变激酶结构域。在表达 FLS2(S938A)或 FLS2(D997A)(激酶催化结构域突变体)的拟南芥转基因植物中,flg22 暴露后 BIK1 和同源物的磷酸化被破坏,但在 FLS2(S938D)植物中正常诱导。BIK1 与 FLS2 的结合需要一个激酶活性的 FLS2,但 FLS2-BAK1 结合不需要。因此,FLS2-BIK1 解离和 FLS2-BAK1 结合不足以激活 FLS2 介导的防御,但是所提出的 FLS2 磷酸化位点 Ser-938 和 FLS2 激酶活性对于整体防御激活以及适当的 flg22 刺激的 BIK1 和同源物的磷酸化都是必需的。