Institut de Biologie Moléculaire des Plantes, Centre National de la Recherche Scientifique (CNRS), Université de Strasbourg, 12 rue du général Zimmer, 67084 Strasbourg Cedex, France.
Nucleic Acids Res. 2013 Aug;41(14):7115-27. doi: 10.1093/nar/gkt465. Epub 2013 Jun 6.
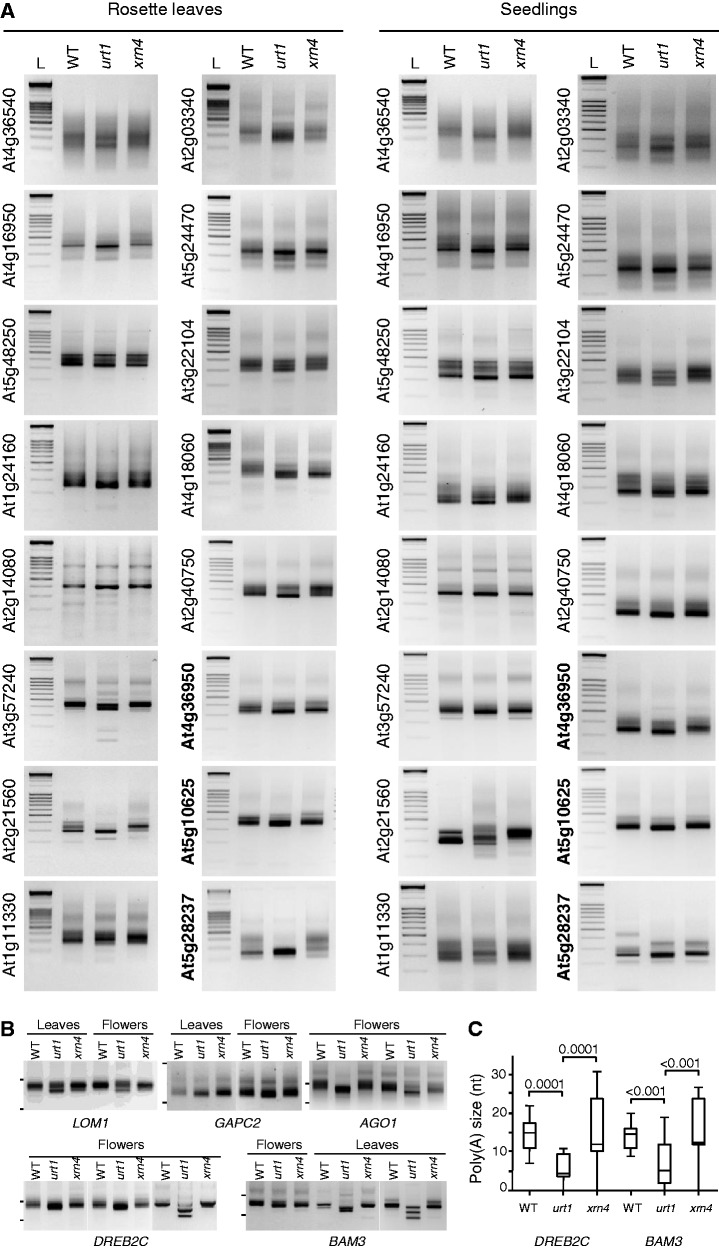
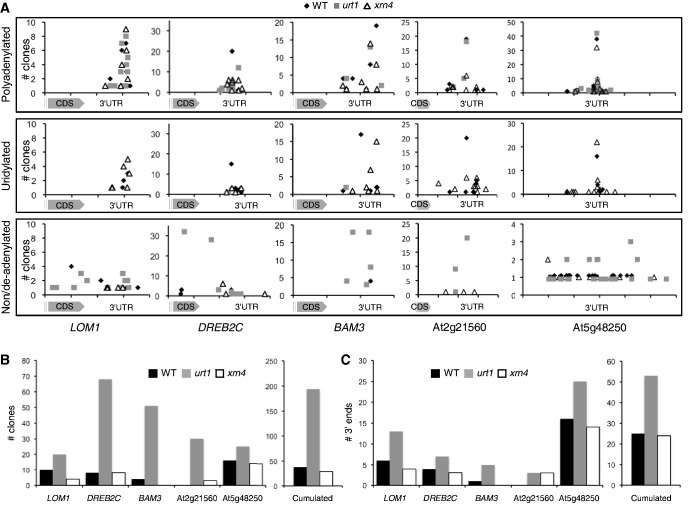
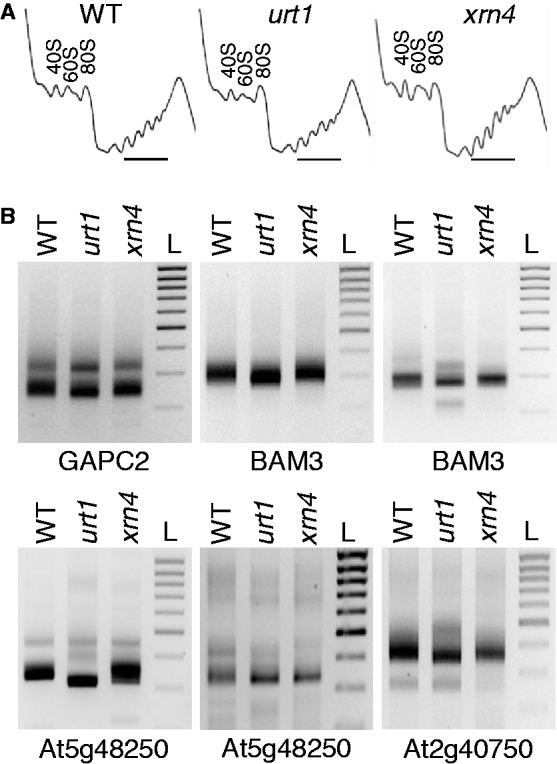
Degradation of mRNAs is usually initiated by deadenylation, the shortening of long poly(A) tails to oligo(A) tails of 12-15 As. Deadenylation leads to decapping and to subsequent 5' to 3' degradation by XRN proteins, or alternatively 3' to 5' degradation by the exosome. Decapping can also be induced by uridylation as shown for the non-polyadenylated histone mRNAs in humans and for several mRNAs in Schizosaccharomyces pombe and Aspergillus nidulans. Here we report a novel role for uridylation in preventing 3' trimming of oligoadenylated mRNAs in Arabidopsis. We show that oligo(A)-tailed mRNAs are uridylated by the cytosolic UTP:RNA uridylyltransferase URT1 and that URT1 has no major impact on mRNA degradation rates. However, in absence of uridylation, oligo(A) tails are trimmed, indicating that uridylation protects oligoadenylated mRNAs from 3' ribonucleolytic attacks. This conclusion is further supported by an increase in 3' truncated transcripts detected in urt1 mutants. We propose that preventing 3' trimming of oligo(A)-tailed mRNAs by uridylation participates in establishing the 5' to 3' directionality of mRNA degradation. Importantly, uridylation prevents 3' shortening of mRNAs associated with polysomes, suggesting that a key biological function of uridylation is to confer 5' to 3' polarity in case of co-translational mRNA decay.
mRNA 的降解通常由去腺苷酸化引发,即长的 poly(A)尾巴缩短为 12-15 个 A 的寡聚(A)尾巴。去腺苷酸化导致脱帽,随后由 XRN 蛋白进行 5'到 3'降解,或者由外切体进行 3'到 5'降解。正如人类非多聚腺苷酸化组蛋白 mRNA 和酿酒酵母和构巢曲霉中的几种 mRNA 所显示的那样,脱帽也可以被尿苷化诱导。在这里,我们报告了尿苷化在防止拟南芥寡聚腺苷酸化 mRNA 3'修剪中的一个新作用。我们表明,胞质 UTP:RNA 尿苷酰转移酶 URT1 将 poly(A)尾巴尿苷化,并且 URT1 对 mRNA 降解速率没有重大影响。然而,在没有尿苷化的情况下,寡聚(A)尾巴被修剪,表明尿苷化保护寡聚腺苷酸化的 mRNA 免受 3'核糖核酸酶攻击。这一结论进一步得到了在 urt1 突变体中检测到的 3'截断转录本增加的支持。我们提出,尿苷化防止寡聚腺苷酸化 mRNA 的 3'修剪参与建立 mRNA 降解的 5'到 3'方向性。重要的是,尿苷化防止与多核糖体相关的 mRNAs 的 3'缩短,这表明尿苷化的一个关键生物学功能是在共翻译 mRNA 降解的情况下赋予 5'到 3'极性。