University of California Los Angeles, David Geffen School of Medicine, Department of Biological Chemistry, Jonsson Comprehensive Cancer Center, Molecular Biology Institute, Bioinformatics Interdepartmental Degree Program, Eli and Edythe Broad Center of Regenerative Medicine and Stem Cell Research, Los Angeles, California 90095, USA.
Nat Cell Biol. 2013 Jul;15(7):872-82. doi: 10.1038/ncb2768. Epub 2013 Jun 9.
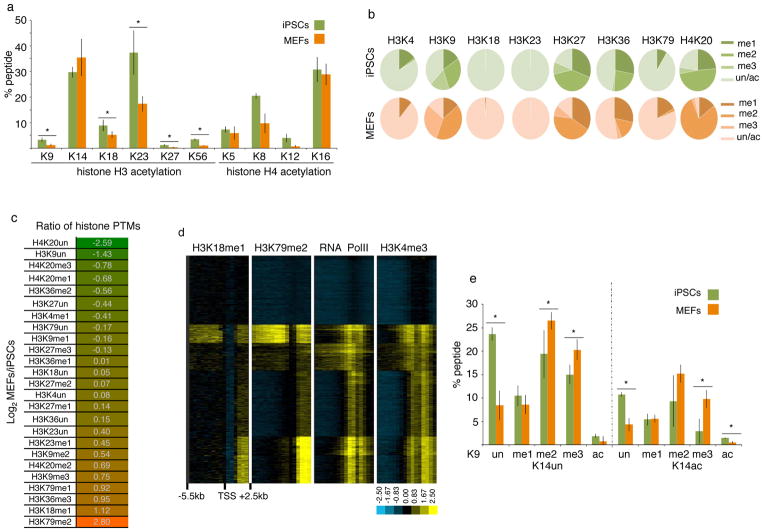
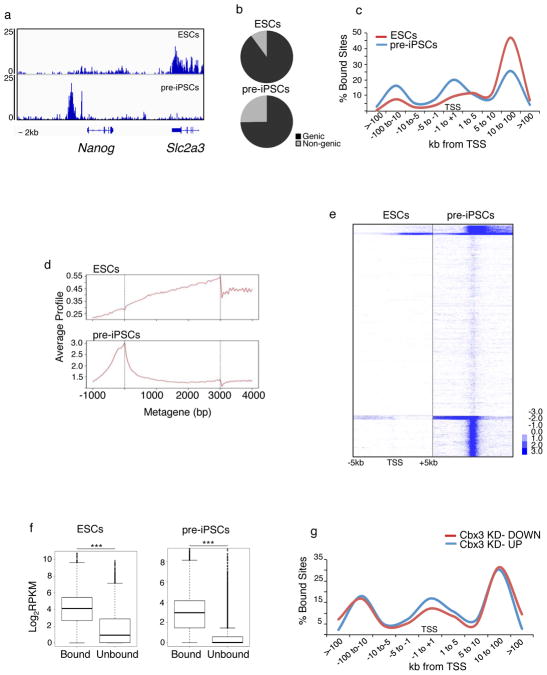
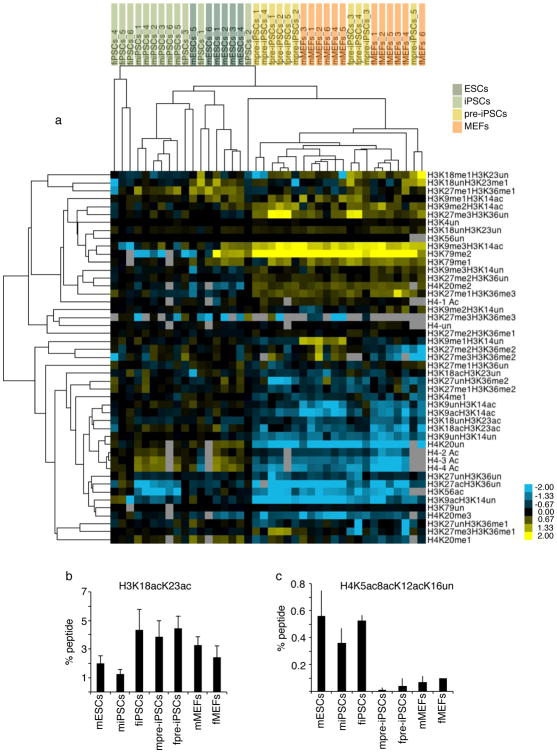
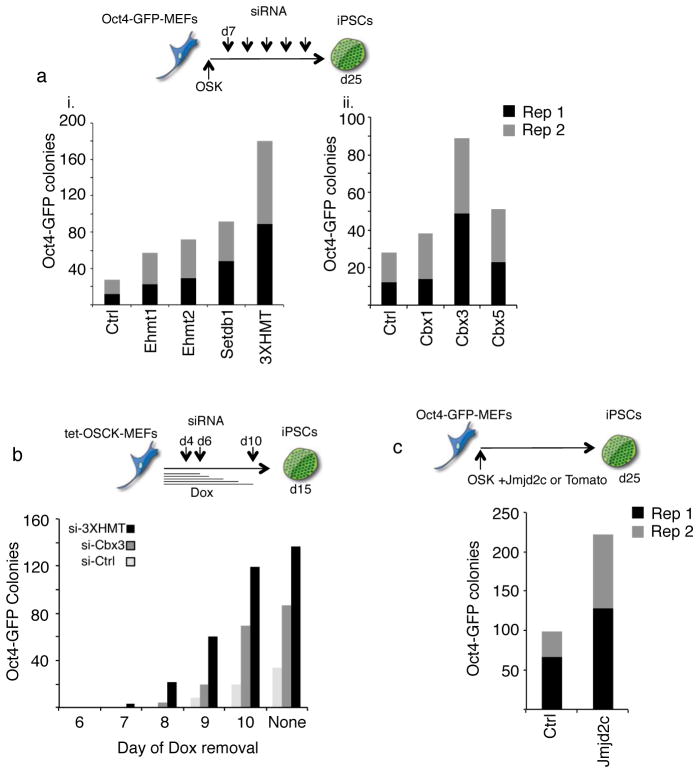
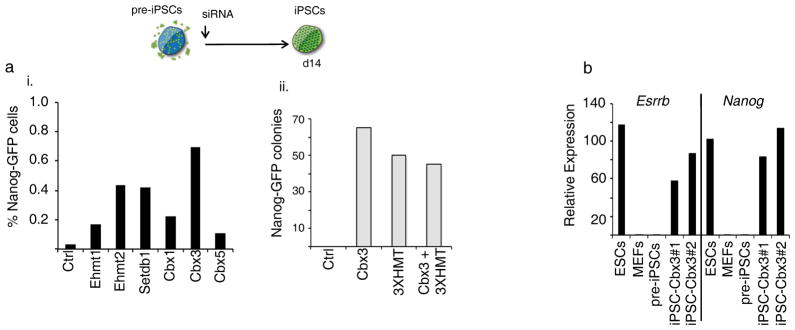
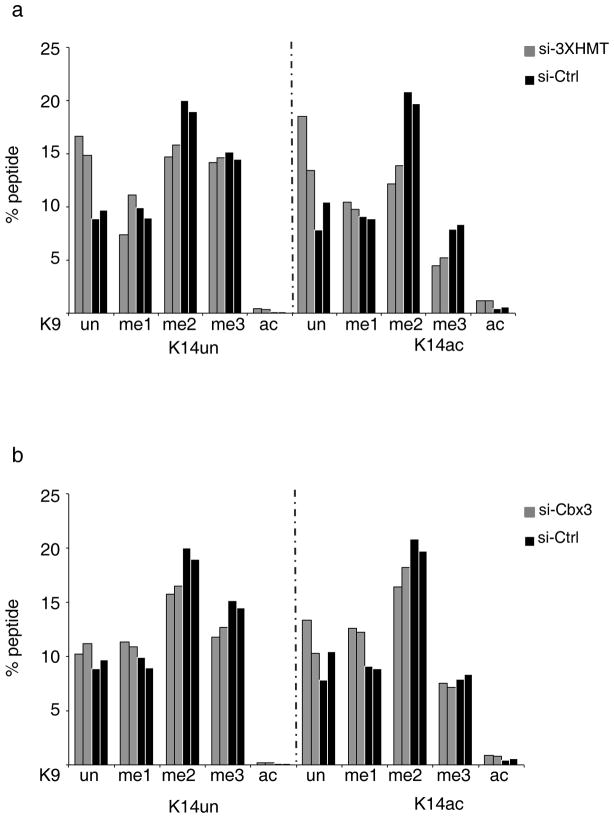
Reprogramming of somatic cells into induced pluripotent stem cells (iPSCs) involves a marked reorganization of chromatin. To identify post-translational histone modifications that change in global abundance during this process, we have applied a quantitative mass-spectrometry-based approach. We found that iPSCs, compared with both the starting fibroblasts and a late reprogramming intermediate (pre-iPSCs), are enriched for histone modifications associated with active chromatin, and depleted for marks of transcriptional elongation and a subset of repressive modifications including H3K9me2/me3. Dissecting the contribution of H3K9 methylation to reprogramming, we show that the H3K9 methyltransferases Ehmt1, Ehmt2 and Setdb1 regulate global H3K9me2/me3 levels and that their depletion increases iPSC formation from both fibroblasts and pre-iPSCs. Similarly, we find that inhibition of heterochromatin protein-1γ (Cbx3), a protein known to recognize H3K9 methylation, enhances reprogramming. Genome-wide location analysis revealed that Cbx3 predominantly binds active genes in both pre-iPSCs and pluripotent cells but with a strikingly different distribution: in pre-iPSCs, but not in embryonic stem cells, Cbx3 associates with active transcriptional start sites, suggesting a developmentally regulated role for Cbx3 in transcriptional activation. Despite largely non-overlapping functions and the predominant association of Cbx3 with active transcription, the H3K9 methyltransferases and Cbx3 both inhibit reprogramming by repressing the pluripotency factor Nanog. Together, our findings demonstrate that Cbx3 and H3K9 methylation restrict late reprogramming events, and suggest that a marked change in global chromatin character constitutes an epigenetic roadblock for reprogramming.
体细胞重编程为诱导多能干细胞(iPSCs)涉及染色质的明显重排。为了鉴定在这个过程中全局丰度发生变化的翻译后组蛋白修饰,我们应用了一种基于定量质谱的方法。我们发现,与起始成纤维细胞和晚期重编程中间产物(pre-iPSCs)相比,iPSCs 富含与活性染色质相关的组蛋白修饰,而缺乏转录延伸的标记和包括 H3K9me2/me3 在内的一组抑制性修饰。剖析 H3K9 甲基化对重编程的贡献,我们表明 H3K9 甲基转移酶 Ehmt1、Ehmt2 和 Setdb1 调节全局 H3K9me2/me3 水平,并且它们的耗竭增加了来自成纤维细胞和 pre-iPSCs 的 iPSC 形成。同样,我们发现抑制异染色质蛋白-1γ(Cbx3),一种已知识别 H3K9 甲基化的蛋白质,可增强重编程。全基因组定位分析表明,Cbx3 主要在 pre-iPSCs 和多能细胞中结合活性基因,但分布明显不同:在 pre-iPSCs 中,但不在胚胎干细胞中,Cbx3 与活性转录起始位点结合,表明 Cbx3 在转录激活中具有发育调控作用。尽管功能主要不重叠,并且 Cbx3 主要与活性转录相关联,但 H3K9 甲基转移酶和 Cbx3 都通过抑制多能因子 Nanog 来抑制重编程。总之,我们的发现表明 Cbx3 和 H3K9 甲基化限制晚期重编程事件,并表明全局染色质特征的显著变化构成了重编程的表观遗传障碍。