Department of Comparative Anatomy and Developmental Biology, Institute of Biology, Faculty of Science, University of Pécs Pécs, Hungary ; Neurobiology and Genetics, Biocenter, Theodor-Boveri-Institute, University of Würzburg Würzburg, Germany.
Front Neural Circuits. 2013 Aug 1;7:127. doi: 10.3389/fncir.2013.00127. eCollection 2013.
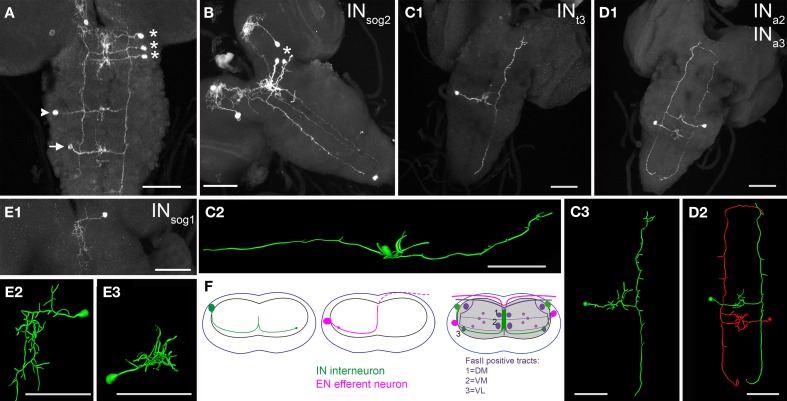
Peptidergic neurons are not easily integrated into current connectomics concepts, since their peptide messages can be distributed via non-synaptic paracrine signaling or volume transmission. Moreover, the polarity of peptidergic interneurons in terms of in- and out-put sites can be hard to predict and is very little explored. We describe in detail the morphology and the subcellular distribution of fluorescent vesicle/dendrite markers in CCAP neurons (NCCAP), a well defined set of peptidergic neurons in the Drosophila larva. NCCAP can be divided into five morphologically distinct subsets. In contrast to other subsets, serial homologous interneurons in the ventral ganglion show a mixed localization of in- and output markers along ventral neurites that defy a classification as dendritic or axonal compartments. Ultrastructurally, these neurites contain both pre- and postsynaptic sites preferably at varicosities. A significant portion of the synaptic events are due to reciprocal synapses. Peptides are mostly non-synaptically or parasynaptically released, and dense-core vesicles and synaptic vesicle pools are typically well separated. The responsiveness of the NCCAP to ecdysis-triggering hormone may be at least partly dependent on a tonic synaptic inhibition, and is independent of ecdysteroids. Our results reveal a remarkable variety and complexity of local synaptic circuitry within a chemically defined set of peptidergic neurons. Synaptic transmitter signaling as well as peptidergic paracrine signaling and volume transmission from varicosities can be main signaling modes of peptidergic interneurons depending on the subcellular region. The possibility of region-specific variable signaling modes should be taken into account in connectomic studies that aim to dissect the circuitry underlying insect behavior and physiology, in which peptidergic neurons act as important regulators.
肽能神经元不易纳入当前的连接组学概念,因为它们的肽信息可以通过非突触旁分泌信号或容积传递来传递。此外,肽能中间神经元在输入和输出部位的极性很难预测,并且很少被探索。我们详细描述了在果蝇幼虫中被定义明确的一组肽能神经元 CCAP 神经元(NCCAP)中荧光囊泡/树突标记物的形态和亚细胞分布。NCCAP 可以分为五个形态上明显不同的亚群。与其他亚群相比,腹神经节中的同源中间神经元沿腹神经纤维显示出输入和输出标记物的混合定位,这使得它们难以分类为树突或轴突区室。超微结构上,这些神经纤维包含优先位于泡状结构处的前突触和后突触位点。一部分突触事件归因于互突触。肽主要通过非突触或旁分泌释放,致密核心囊泡和突触小泡池通常是很好分离的。NCCAP 对蜕皮激素触发激素的反应至少部分取决于持续的突触抑制,并且与蜕皮激素无关。我们的结果揭示了在一组化学定义的肽能神经元中存在着显著的多样性和复杂性的局部突触回路。突触递质信号以及来自泡状结构的肽能旁分泌信号和容积传递可以是肽能中间神经元的主要信号模式,这取决于亚细胞区域。在连接组学研究中,应考虑到特定区域的可变信号模式的可能性,这些研究旨在剖析昆虫行为和生理学背后的电路,其中肽能神经元作为重要的调节剂。