Bioinformatik, Institut für Biochemie, Friedrich-Alexander-Universität Erlangen-Nürnberg, Erlangen, Germany.
PLoS One. 2013 Jul 31;8(7):e70521. doi: 10.1371/journal.pone.0070521. Print 2013.
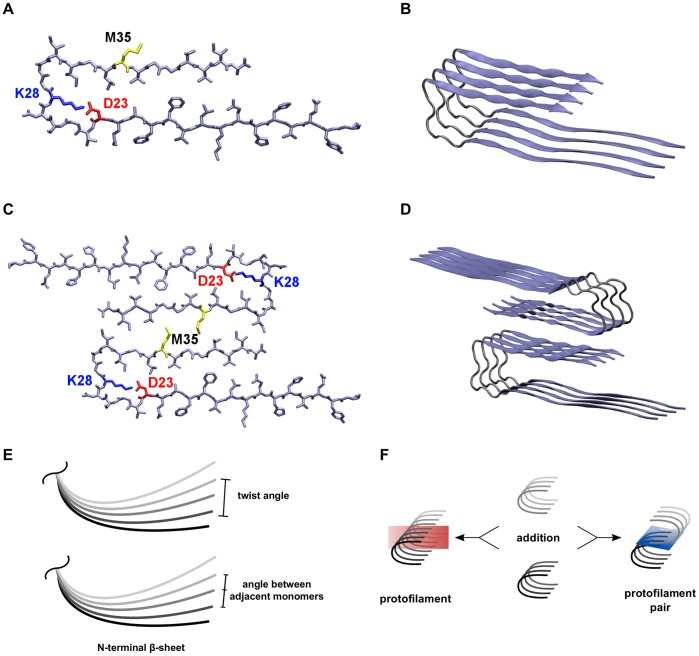
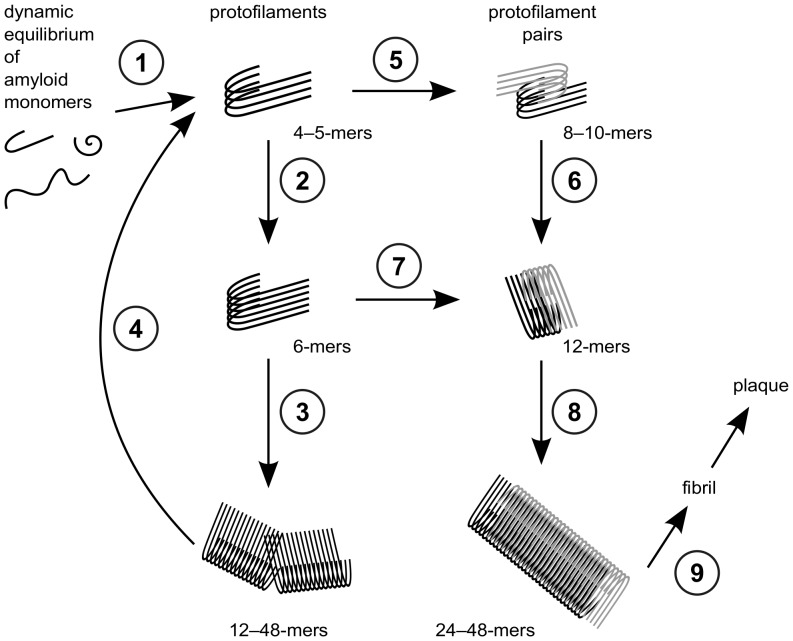
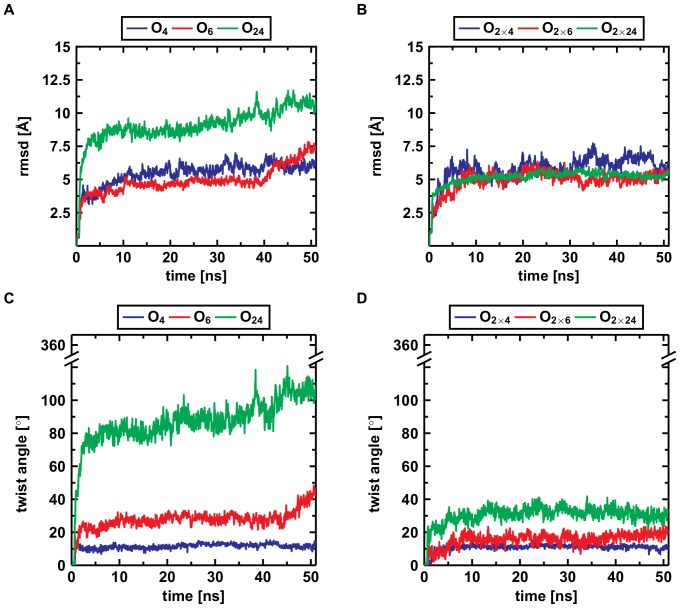
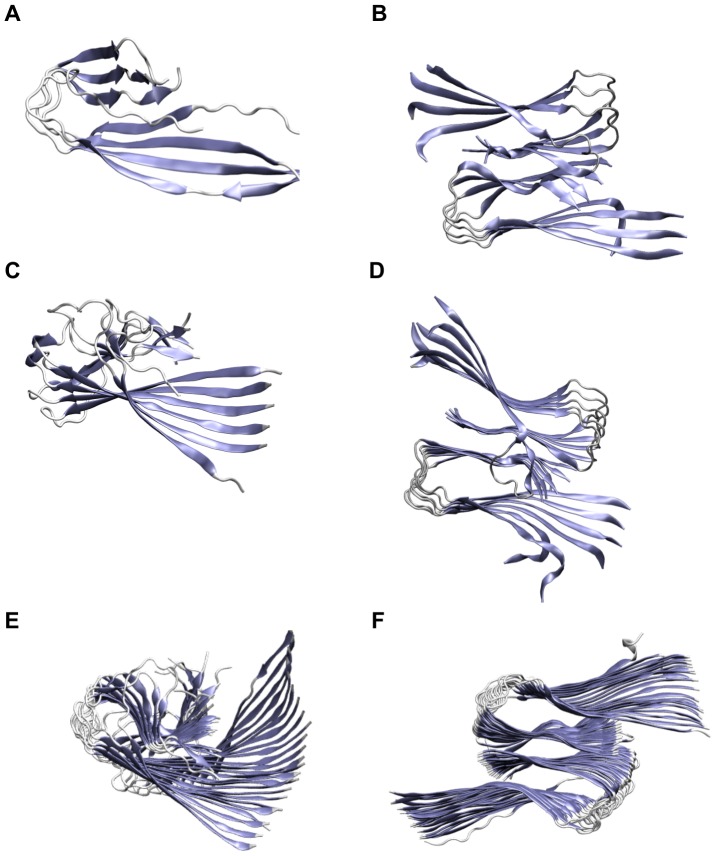
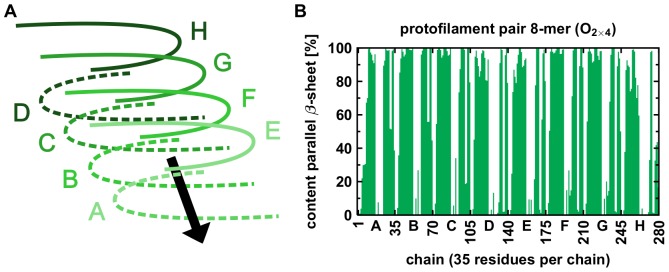
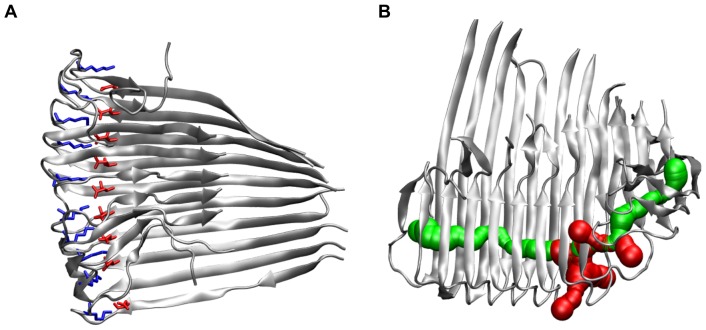
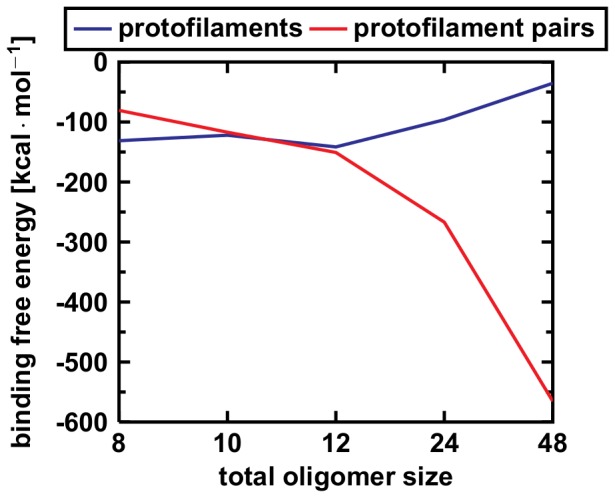
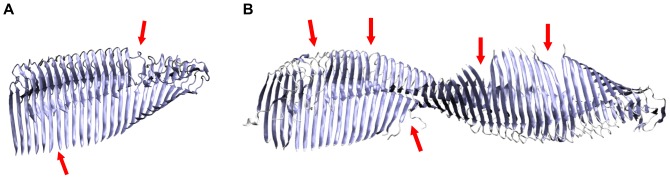
Amyloid-[Formula: see text] (A[Formula: see text]) oligomers play a crucial role in Alzheimer's disease due to their neurotoxic aggregation properties. Fibrillar A[Formula: see text] oligomerization can lead to protofilaments and protofilament pairs via oligomer elongation and oligomer association, respectively. Small fibrillar oligomers adopt the protofilament topology, whereas fibrils contain at least protofilament pairs. To date, the underlying growth mechanism from oligomers to the mature fibril still remains to be elucidated. Here, we performed all-atom molecular dynamics simulations in explicit solvent on single layer-like protofilaments and fibril-like protofilament pairs of different size ranging from the tetramer to the 48-mer. We found that the initial U-shaped topology per monomer is maintained over time in all oligomers. The observed deviations of protofilaments from the starting structure increase significantly with size due to the twisting of the in-register parallel [Formula: see text]-sheets. This twist causes long protofilaments to be unstable and leads to a breakage. Protofilament pairs, which are stabilized by a hydrophobic interface, exhibit more fibril-like properties such as the overall structure and the twist angle. Thus, they can act as stable conformational templates for further fibril growth. Key properties like the twist angle, shape complementarity, and energetics show a size-dependent behavior so that small oligomers favor the protofilament topology, whereas large oligomers favor the protofilament pair topology. The region for this conformational transition is at the size of approximately twelve A[Formula: see text] monomers. From that, we propose the following growth mechanism from A[Formula: see text] oligomers to fibrils: (1) elongation of short protofilaments; (2) breakage of large protofilaments; (3) formation of short protofilament pairs; and (4) elongation of protofilament pairs.
淀粉样蛋白-β(Aβ)寡聚体由于其神经毒性聚集特性在阿尔茨海默病中起着至关重要的作用。纤维状 Aβ寡聚体通过寡聚体伸长和寡聚体缔合分别导致原纤维和原纤维对。小纤维状寡聚体采用原纤维拓扑结构,而纤维包含至少原纤维对。迄今为止,寡聚体向成熟纤维的潜在生长机制仍有待阐明。在这里,我们在明溶剂中对不同大小的单层样原纤维和纤维样原纤维对进行了全原子分子动力学模拟,大小范围从四聚体到 48 聚体。我们发现,每个单体的初始 U 形拓扑结构随时间保持不变。由于同序平行β-折叠的扭转,观察到的原纤维偏离起始结构的偏差随尺寸显著增加。这种扭曲使长原纤维不稳定并导致断裂。由疏水性界面稳定的原纤维对表现出更多的纤维状特性,如整体结构和扭转角。因此,它们可以作为进一步纤维生长的稳定构象模板。关键性质,如扭转角、形状互补性和能量学,表现出尺寸依赖性行为,使得小寡聚体有利于原纤维拓扑结构,而大寡聚体有利于原纤维对拓扑结构。这种构象转变的区域在大约十二个 Aβ单体的大小。由此,我们提出了从 Aβ寡聚体到纤维的以下生长机制:(1)短原纤维的伸长;(2)大原纤维的断裂;(3)短原纤维对的形成;和(4)原纤维对的伸长。