Takahashi Yui, Kawabata Hiroaki, Murakami Shuichiro
Department of Agricultural Chemistry, Graduate School of Agriculture, Meiji University, 1-1-1, Higashimita, Tama-ku, Kawasaki, 214-8571 Japan.
Springerplus. 2013 Sep 9;2:447. doi: 10.1186/2193-1801-2-447. eCollection 2013.
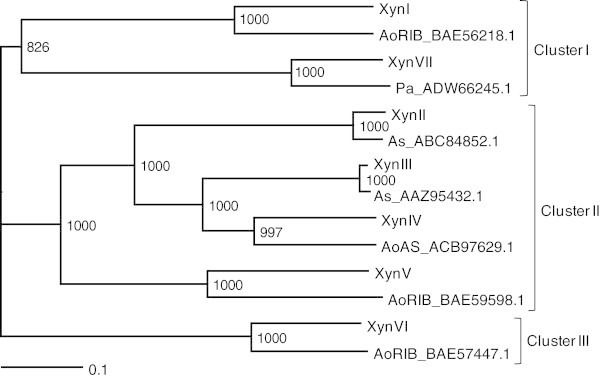
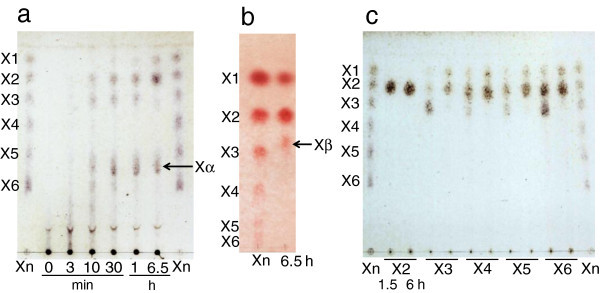
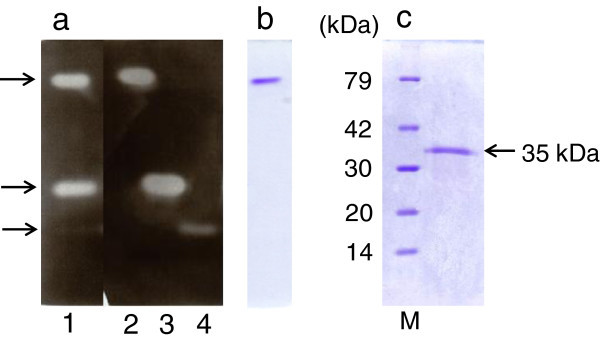
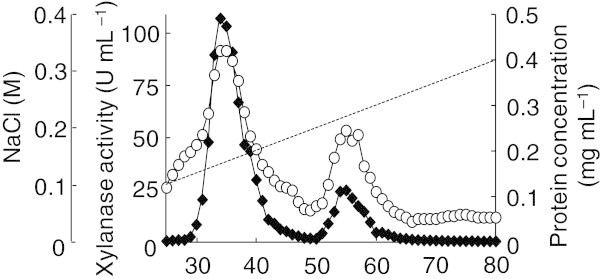
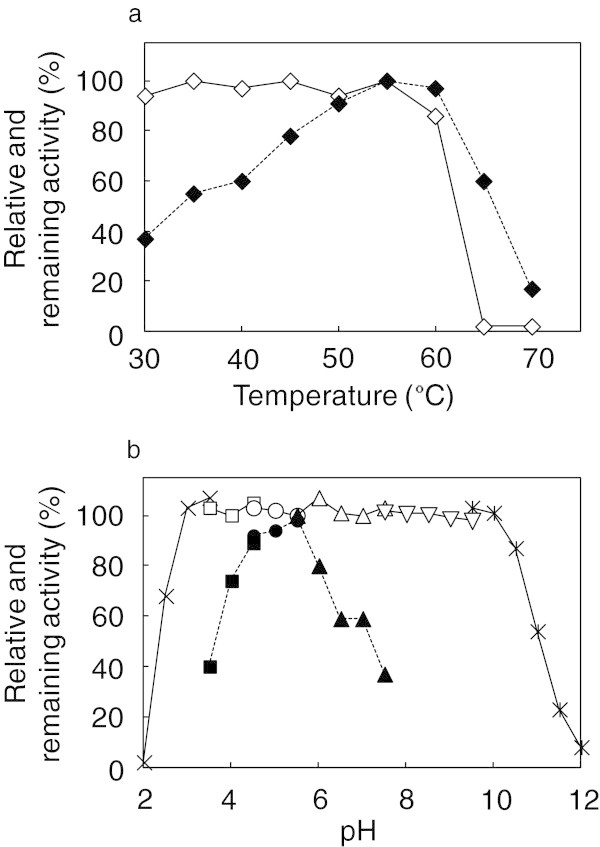
Xylanases produced by Aspergillus niger are industrially important and many types of xylanases have been reported. Individual xylanases have been well studied for their enzymatic properties, gene cloning, and heterologous expression. However, less attention has been paid to the relationship between xylanase genes carried on the A. niger genome and xylanases produced by A. niger strains. Therefore, we examined xylanase genes encoded on the genome of A. niger E-1 and xylanases produced in culture. Seven putative xylanase genes, xynI-VII (named in ascending order of the molecular masses of the deduced amino acid sequences), were amplified from the strain E-1 genome using primers designed from the genome sequence of A. niger CBS 513.88 by PCR and phylogenetically classified into three clusters. Additionally, culture supernatant analysis by DE52 anion-exchange column chromatography revealed that this strain produced three xylanases, XynII, XynIII, and XynVII, which were identified by N-terminal amino acid sequencing and MALDI-TOF-MS analyses, in culture when gown in 0.5% xylan medium supplemented with 50 mM succinate. Furthermore, XynVII, the only GH family 10 xylanase in A. niger E-1, was purified and characterized. The purified enzyme showed a single band with a molecular mass of 35 kDa by SDS-PAGE. The highest activity of purified XynVII was observed at 55°C and pH 5.5. The enzyme was stable in the broad pH range of 3-10 and up to 60°C and was resistant to most metal ions and modifying regents. XynVII showed high specificity against beechwood xylan with K m and V max values of 2.8 mg mL(-1) and 127 μmol min(-1)mg(-1), respectively. TLC and MALDI-TOF-MS analyses showed that the final hydrolyzed products of the enzyme from beechwood xylan were xylose, xylobiose, and xylotriose substituted with a 4-o-metylglucuronic acid residue.
黑曲霉产生的木聚糖酶在工业上具有重要意义,并且已经报道了多种类型的木聚糖酶。单个木聚糖酶的酶学性质、基因克隆和异源表达已得到充分研究。然而,对于黑曲霉基因组中携带的木聚糖酶基因与黑曲霉菌株产生的木聚糖酶之间的关系关注较少。因此,我们研究了黑曲霉E-1基因组中编码的木聚糖酶基因以及培养物中产生的木聚糖酶。使用根据黑曲霉CBS 513.88的基因组序列设计的引物,通过PCR从菌株E-1基因组中扩增出7个推定的木聚糖酶基因,即xynI-VII(按照推导氨基酸序列分子量升序命名),并在系统发育上分为三个簇。此外,通过DE52阴离子交换柱色谱对培养上清液进行分析,结果表明该菌株在添加了50 mM琥珀酸盐的0.5%木聚糖培养基中培养时,在培养物中产生了三种木聚糖酶,即XynII、XynIII和XynVII,通过N端氨基酸测序和基质辅助激光解吸电离飞行时间质谱(MALDI-TOF-MS)分析对其进行了鉴定。此外,对黑曲霉E-1中唯一的糖基水解酶家族10(GH family 10)木聚糖酶XynVII进行了纯化和表征。通过十二烷基硫酸钠聚丙烯酰胺凝胶电泳(SDS-PAGE)分析,纯化后的酶呈现出一条分子量为35 kDa的条带。纯化后的XynVII在55°C和pH 5.5时表现出最高活性。该酶在3-10的较宽pH范围内以及高达60°C时都很稳定,并且对大多数金属离子和修饰剂具有抗性。XynVII对山毛榉木聚糖具有高度特异性,其米氏常数(K m)和最大反应速度(V max)值分别为2.8 mg mL(-1)和12 μmol min(-1)mg(-1)。薄层色谱(TLC)和MALDI-TOF-MS分析表明,该酶对山毛榉木聚糖的最终水解产物是被4-O-甲基葡萄糖醛酸残基取代的木糖、木二糖和木三糖。