Department of Medical Sciences, University of Turin, Turin, Italy.
J Am Heart Assoc. 2013 Dec 5;2(6):e000376. doi: 10.1161/JAHA.113.000376.
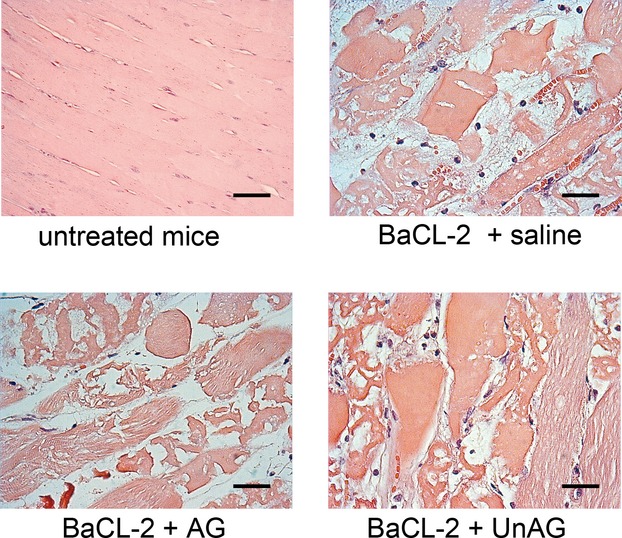
Surgical treatment of peripheral artery disease, even if successful, does not prevent reoccurrence. Under these conditions, increased oxidative stress is a crucial determinant of tissue damage. Given its reported antioxidant effects, we investigated the potential of unacylated-ghrelin (UnAG) to reduce ischemia-induced tissue damage in a mouse model of peripheral artery disease.
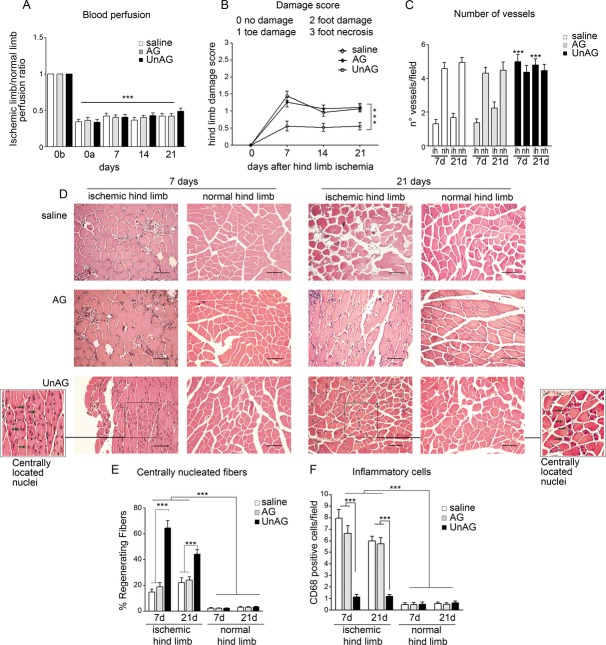
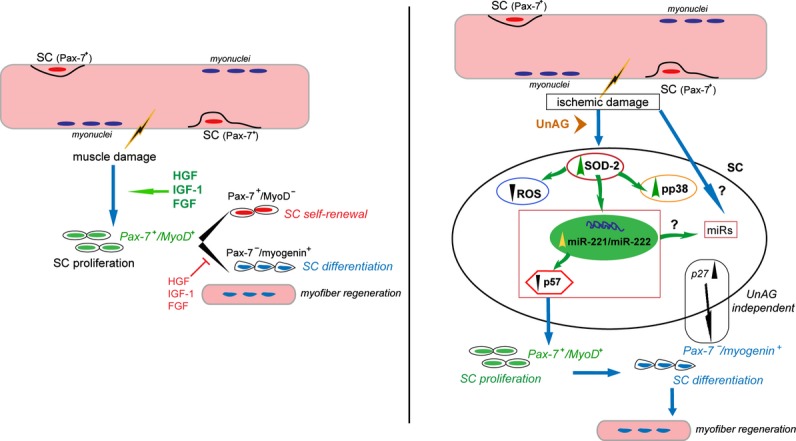
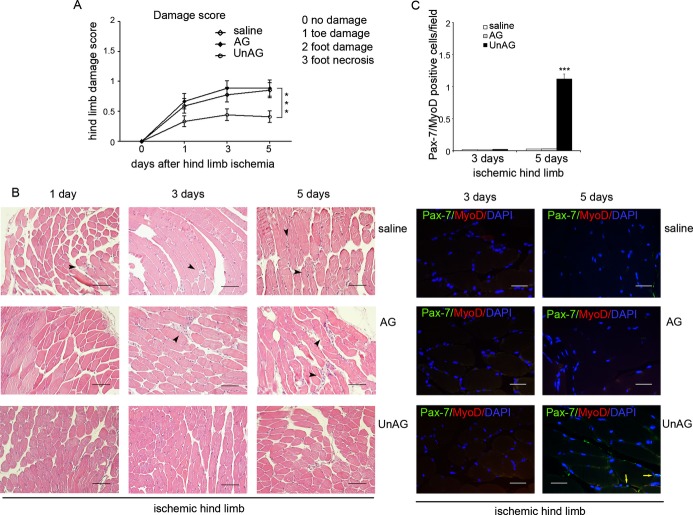
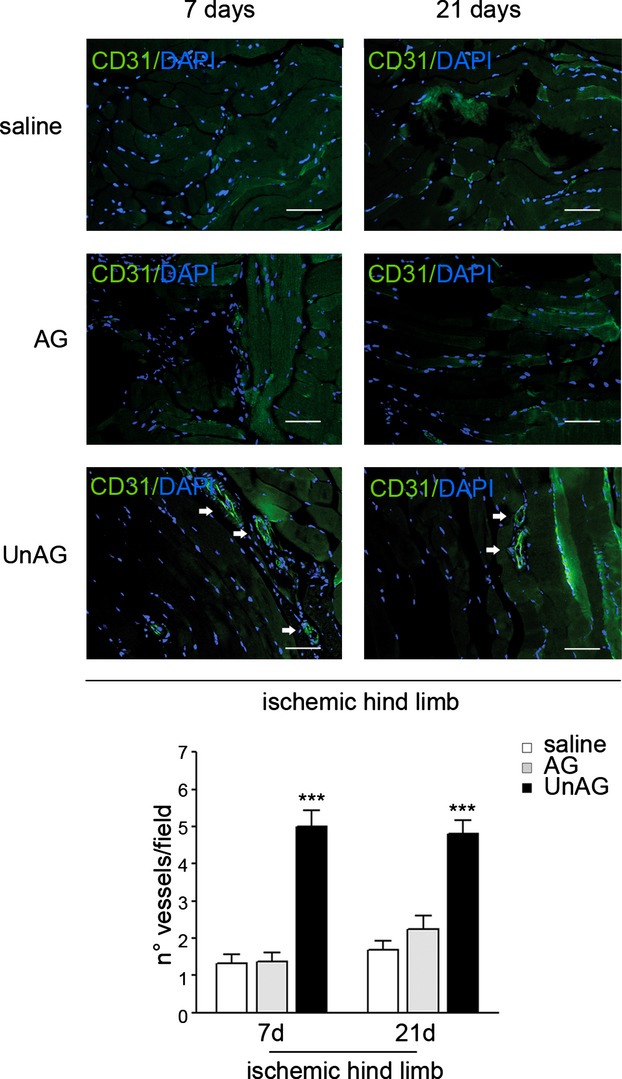
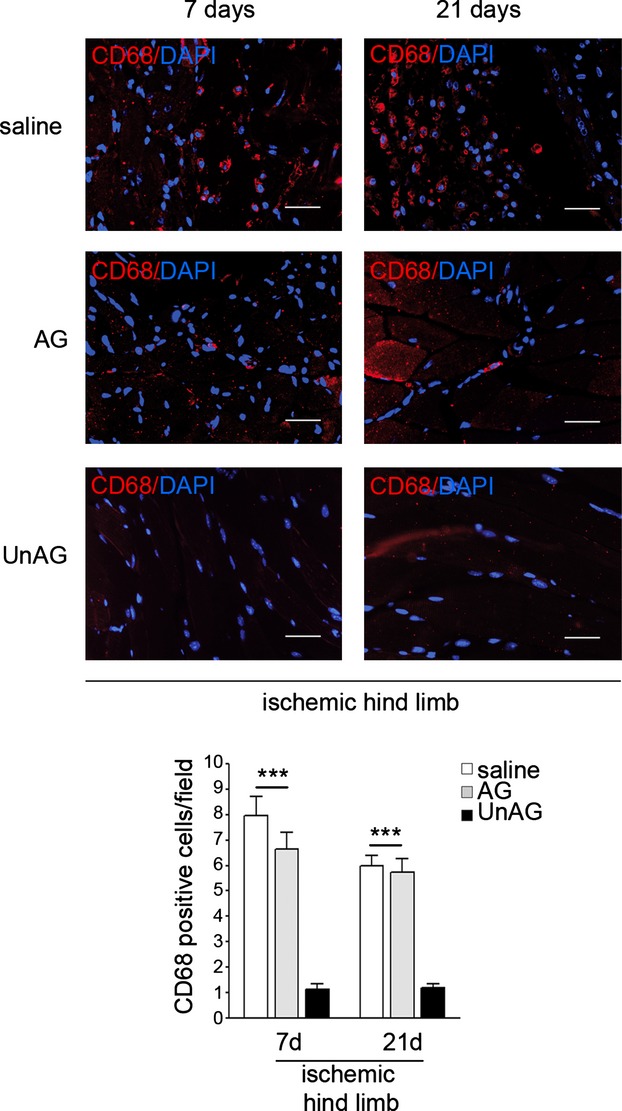
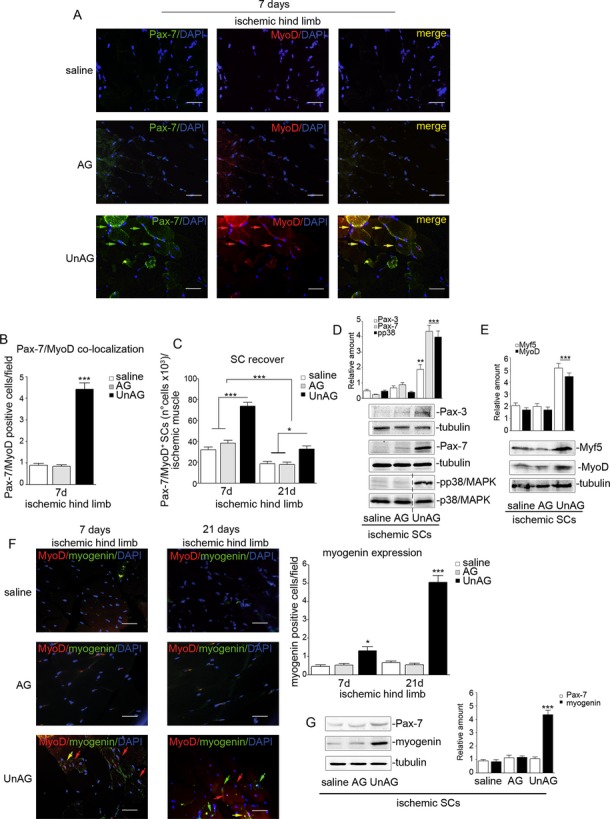
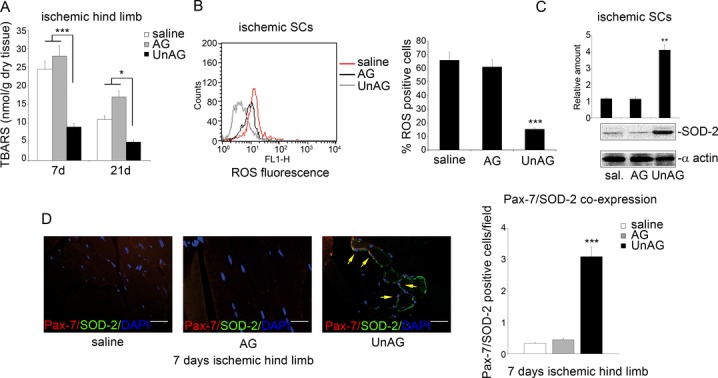
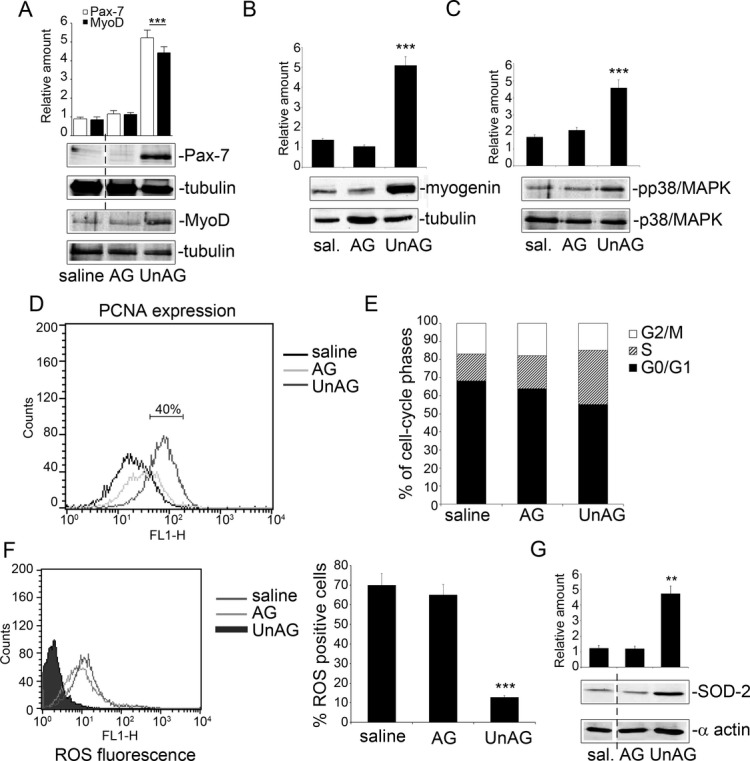
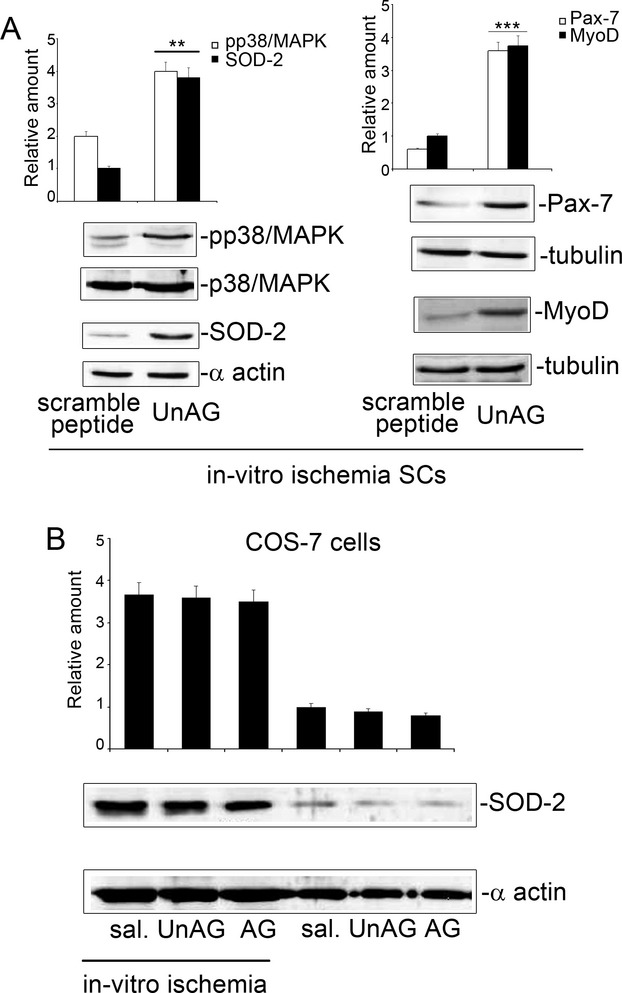
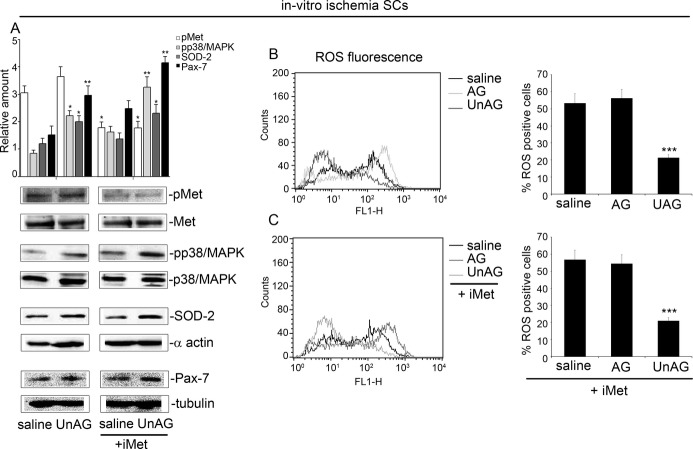
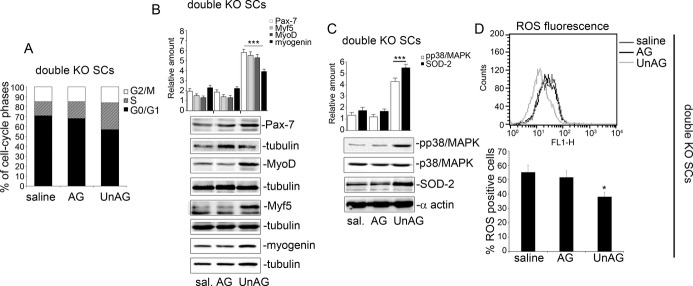
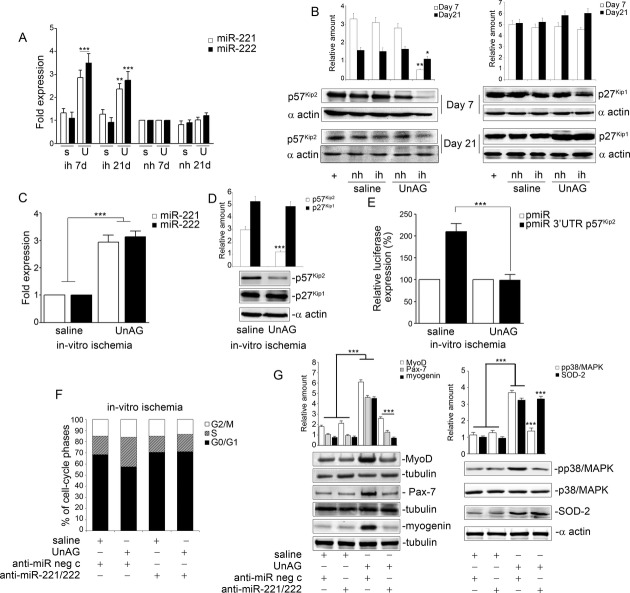
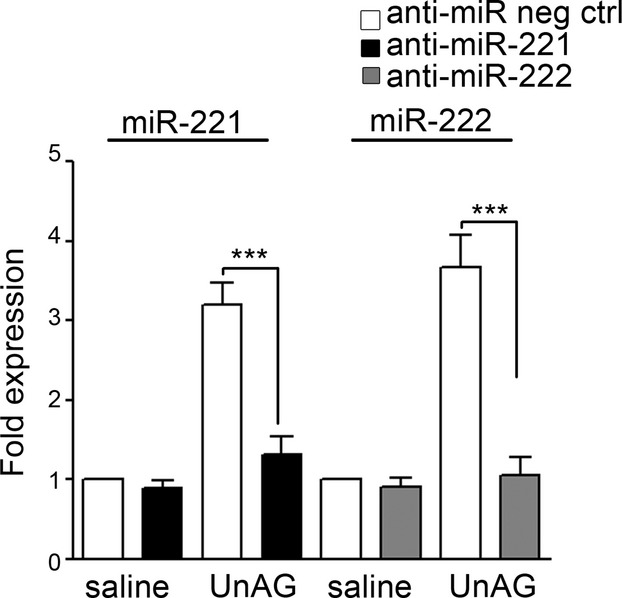
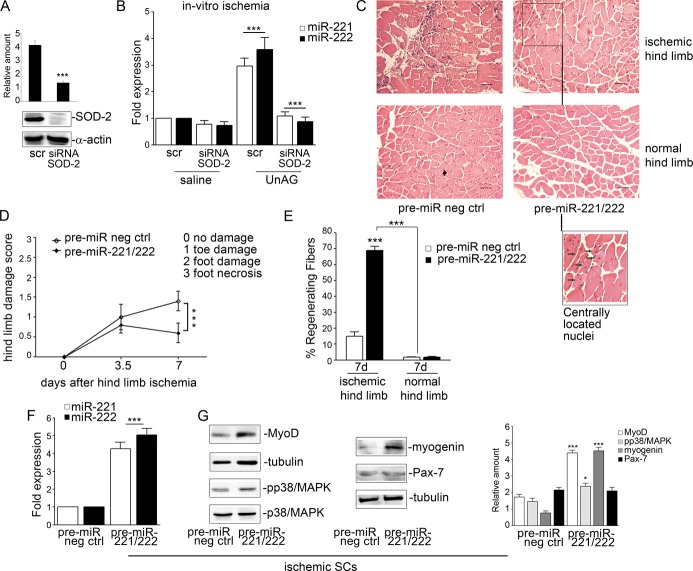
We show that UnAG but not acylated ghrelin (AG) induces skeletal muscle regeneration in response to ischemia via canonical p38/mitogen-actived protein kinase signaling UnAG protected against reactive oxygen species-induced cell injuries by inducing the expression of superoxide dismutase-2 (SOD-2) in satellite cells. This led to a reduced number of infiltrating CD68(+) cells and was followed by induction of the myogenic process and a reduction in functional impairment. Moreover, we found that miR-221/222, previously linked to muscle regeneration processes, was up-regulated and negatively correlated with p57(Kip2) expression in UnAG-treated mice. UnAG, unlike AG, promoted cell-cycle entry in satellite cells of mice lacking the genes for ghrelin and its receptor (GHSR1a). UnAG-induced p38/mitogen-actived protein kinase phosphorylation, leading to activation of the myogenic process, was prevented in SOD-2-depleted SCs. By siRNA technology, we also demonstrated that SOD-2 is the antioxidant enzyme involved in the control of miR-221/222-driven posttranscriptional p57(Kip2) regulation. Loss-of-function experiments targeting miR-221/222 and local pre-miR-221/222 injection in vivo confirmed a role for miR-221/222 in driving skeletal muscle regeneration after ischemia.
These results indicate that UnAG-induced skeletal muscle regeneration after ischemia depends on SOD-2-induced miR-221/222 expression and highlight its clinical potential for the treatment of reactive oxygen species-mediated skeletal muscle damage.
外周动脉疾病的手术治疗,即使成功,也不能预防其复发。在这种情况下,氧化应激的增加是组织损伤的关键决定因素。考虑到其报道的抗氧化作用,我们研究了未酰化 ghrelin(UnAG)在小鼠外周动脉疾病模型中减少缺血引起的组织损伤的潜力。
我们表明,UnAG 而不是酰化 ghrelin(AG)通过经典的 p38/丝裂原激活蛋白激酶信号通路诱导缺血后的骨骼肌再生。UnAG 通过诱导卫星细胞中超氧化物歧化酶 2(SOD-2)的表达来防止活性氧诱导的细胞损伤。这导致浸润的 CD68(+)细胞数量减少,并随后诱导肌生成过程和功能障碍减少。此外,我们发现,先前与肌肉再生过程相关的 miR-221/222 在 UnAG 处理的小鼠中上调,并与 p57(Kip2)的表达呈负相关。与 AG 不同,UnAG 促进了缺乏 ghrelin 和其受体(GHSR1a)基因的小鼠卫星细胞的细胞周期进入。UnAG 诱导的 p38/丝裂原激活蛋白激酶磷酸化,导致肌生成过程的激活,在 SOD-2 耗尽的 SCs 中被阻止。通过 siRNA 技术,我们还证明了 SOD-2 是控制 miR-221/222 驱动的 p57(Kip2)转录后调控的抗氧化酶。针对 miR-221/222 的功能丧失实验和体内局部 pre-miR-221/222 注射证实了 miR-221/222 在缺血后驱动骨骼肌再生中的作用。
这些结果表明,UnAG 诱导的缺血后骨骼肌再生依赖于 SOD-2 诱导的 miR-221/222 表达,并强调了其在治疗活性氧介导的骨骼肌损伤方面的临床潜力。