Williams Byron C, Filter Joshua J, Blake-Hodek Kristina A, Wadzinski Brian E, Fuda Nicholas J, Shalloway David, Goldberg Michael L
Department of Molecular Biology and Genetics, Cornell University, Ithaca, United States.
Elife. 2014 Mar 11;3:e01695. doi: 10.7554/eLife.01695.
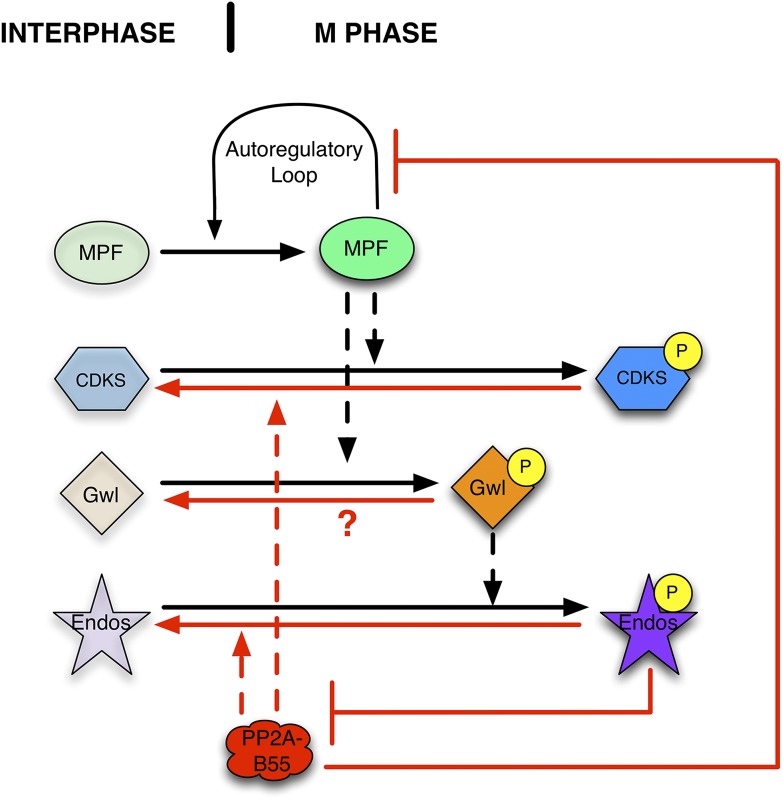
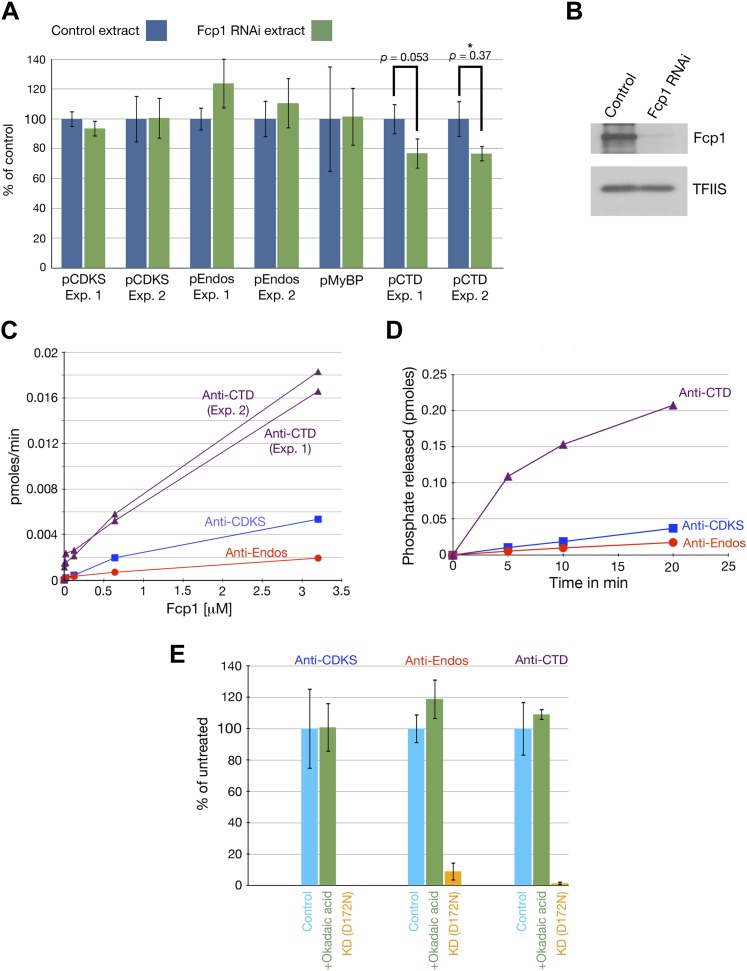
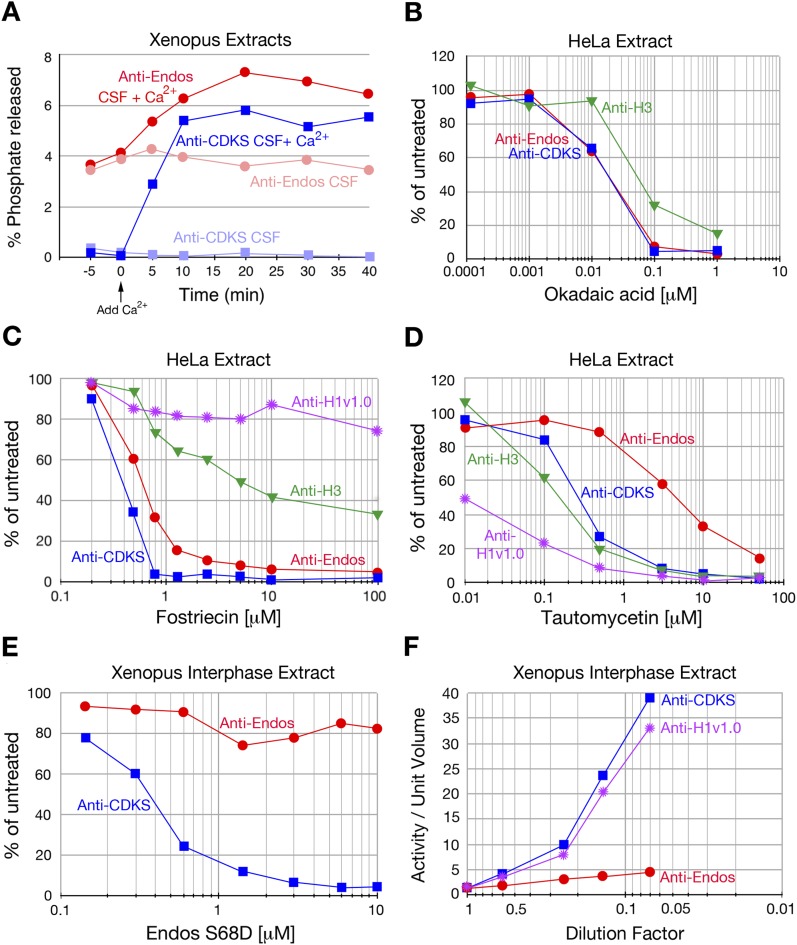
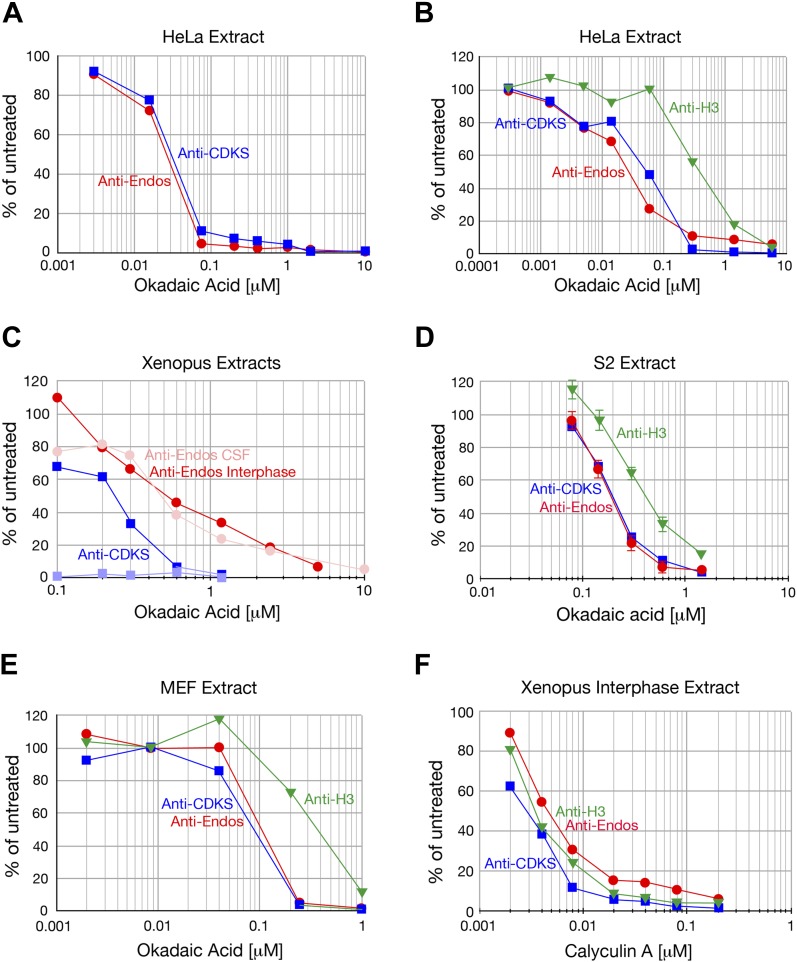
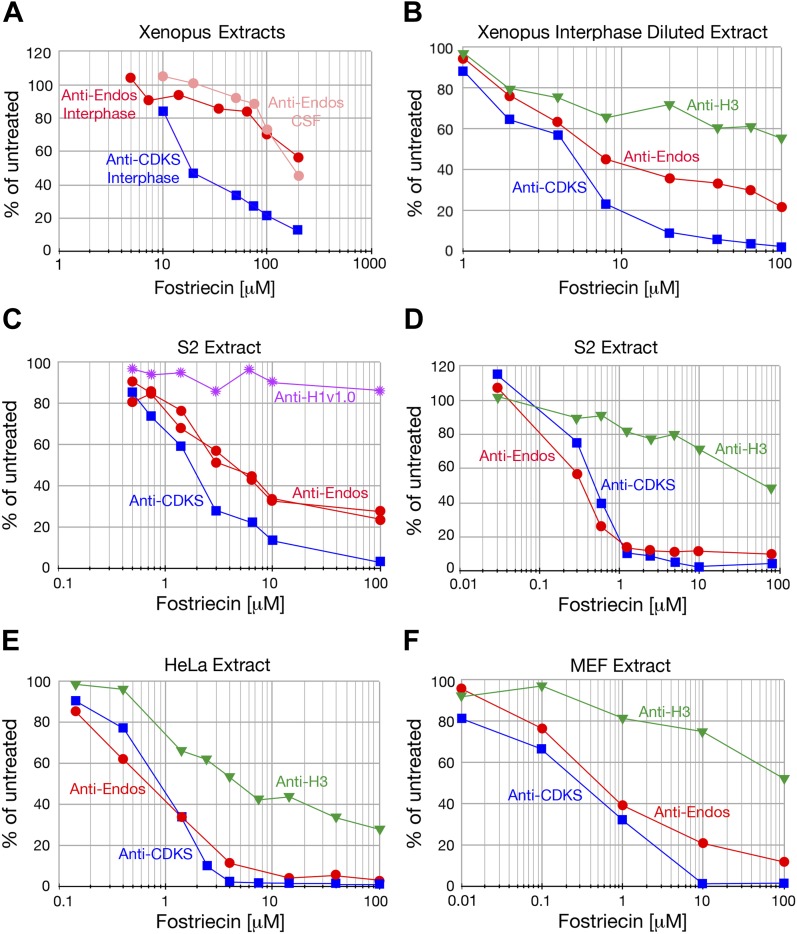
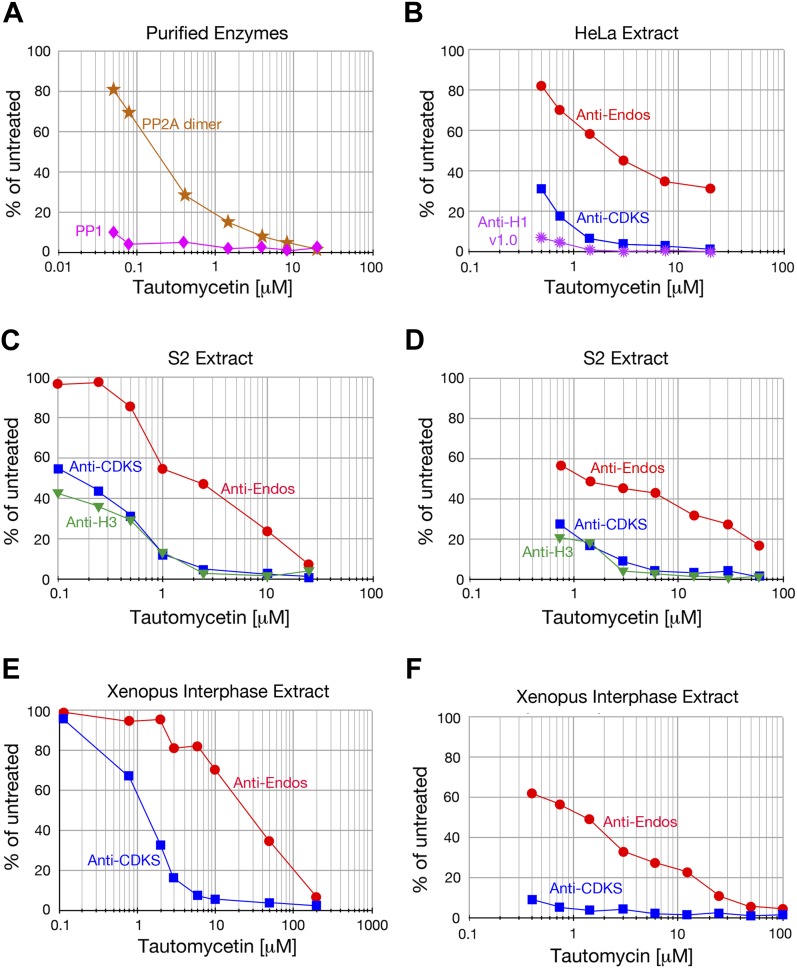
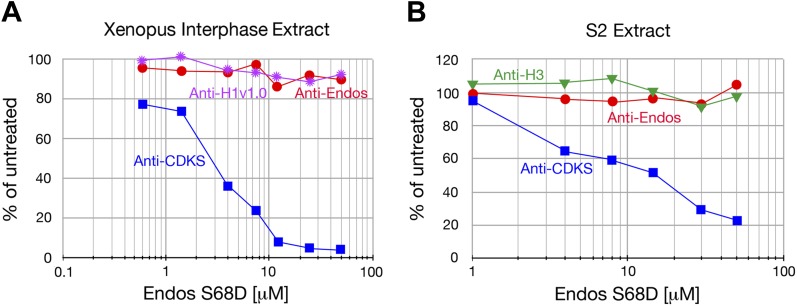
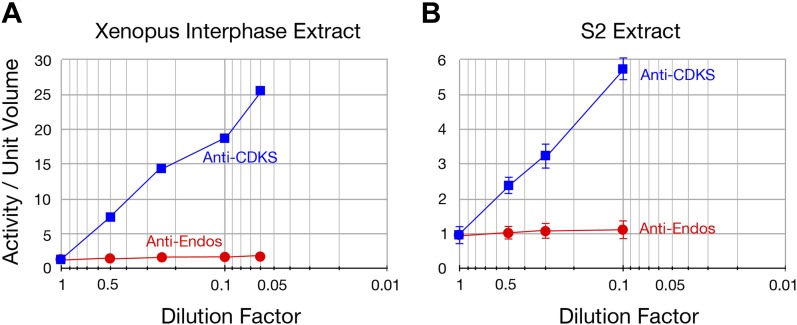
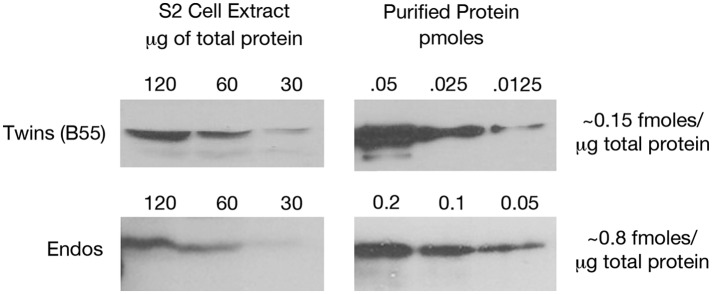
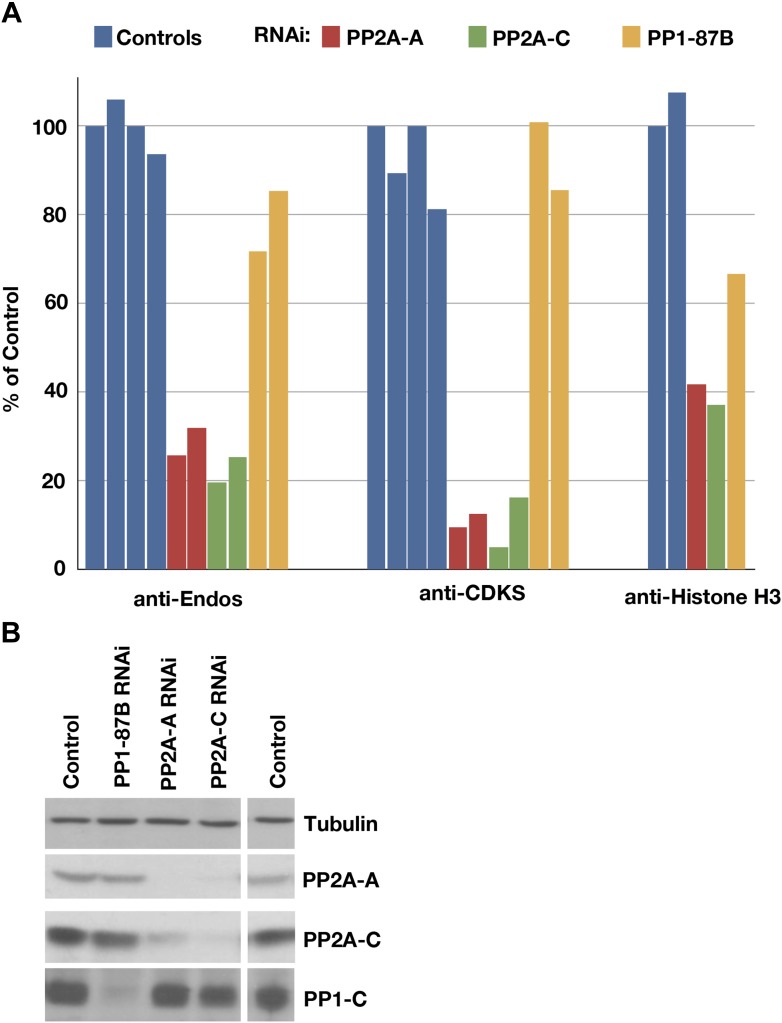
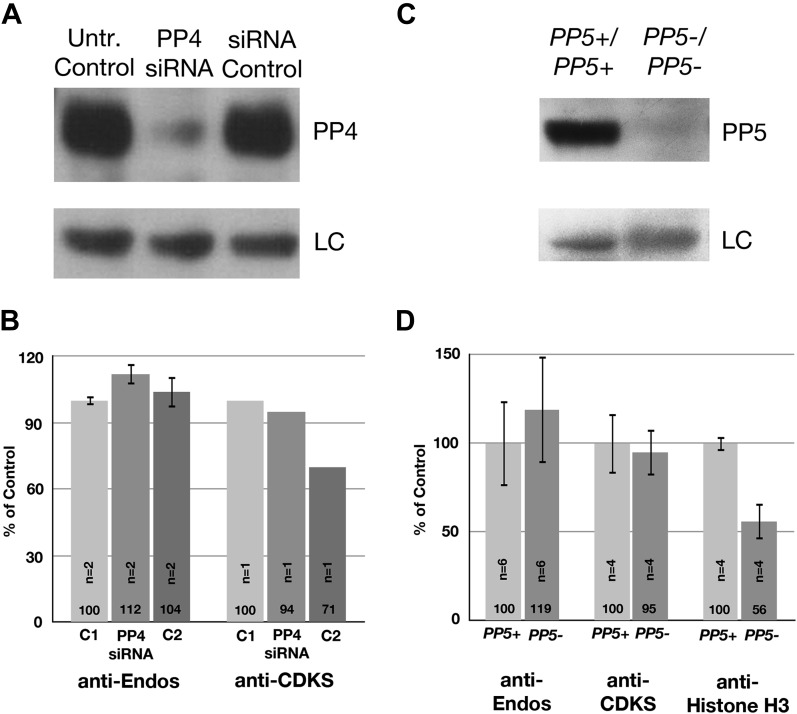
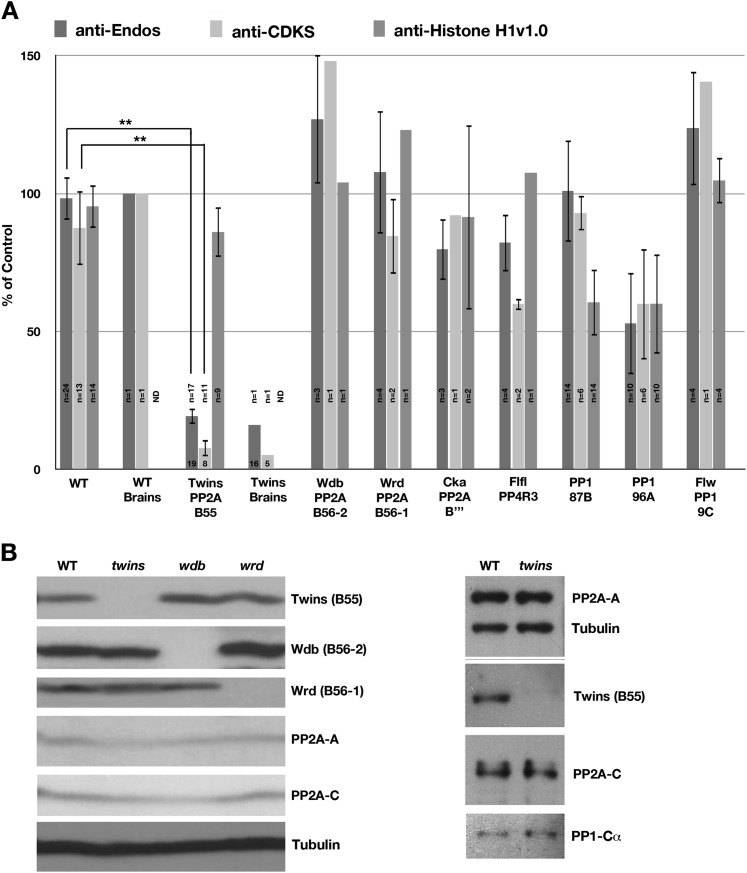
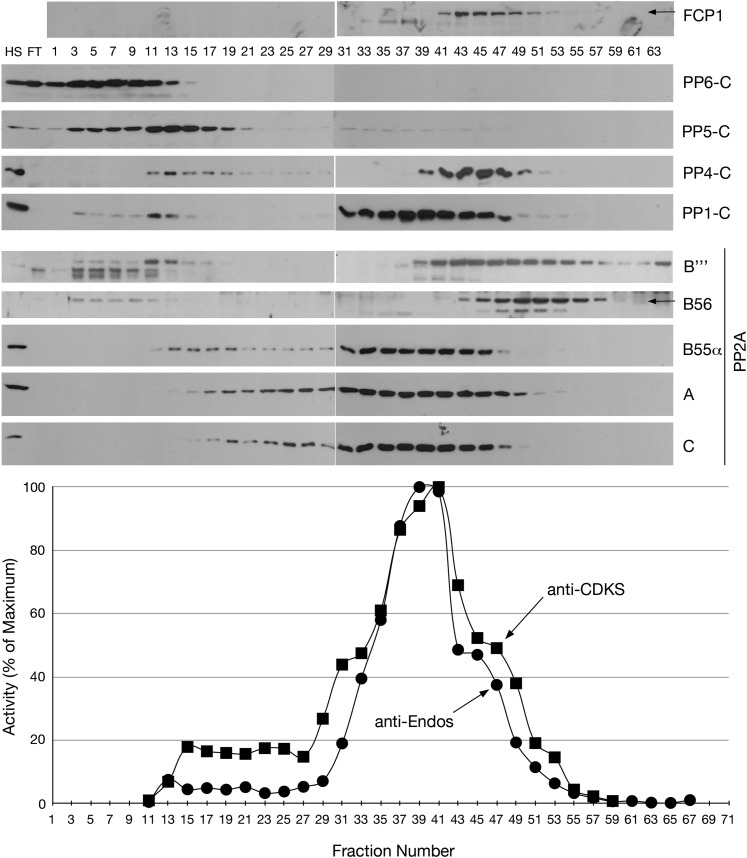
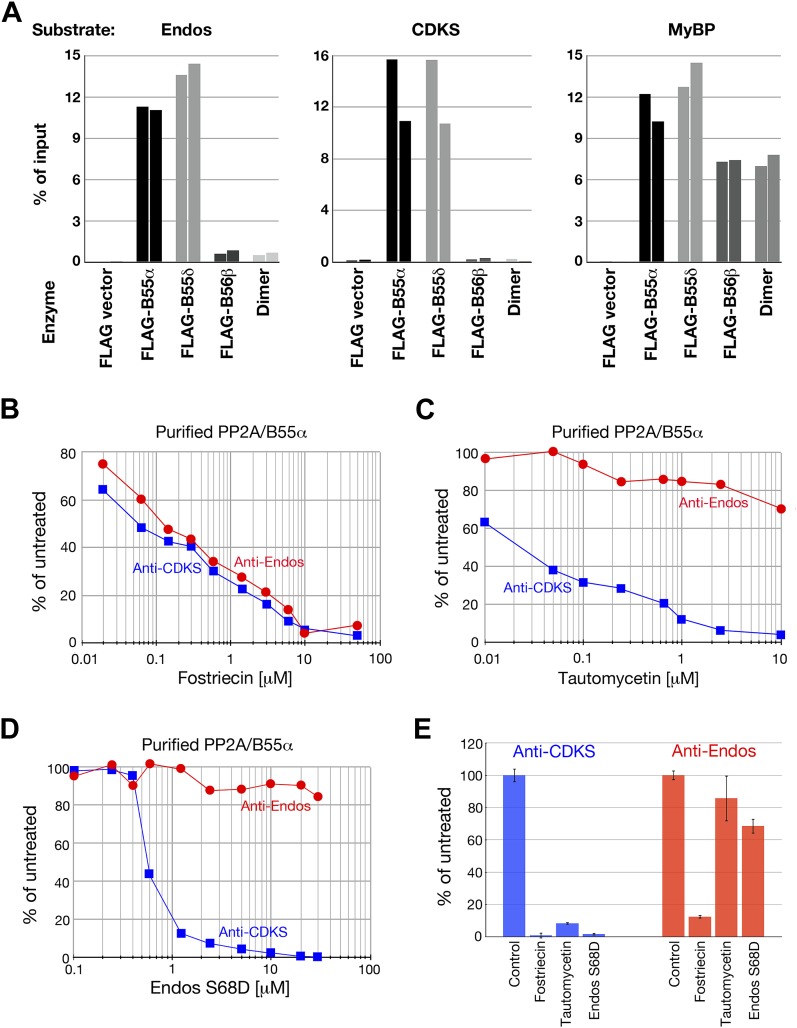
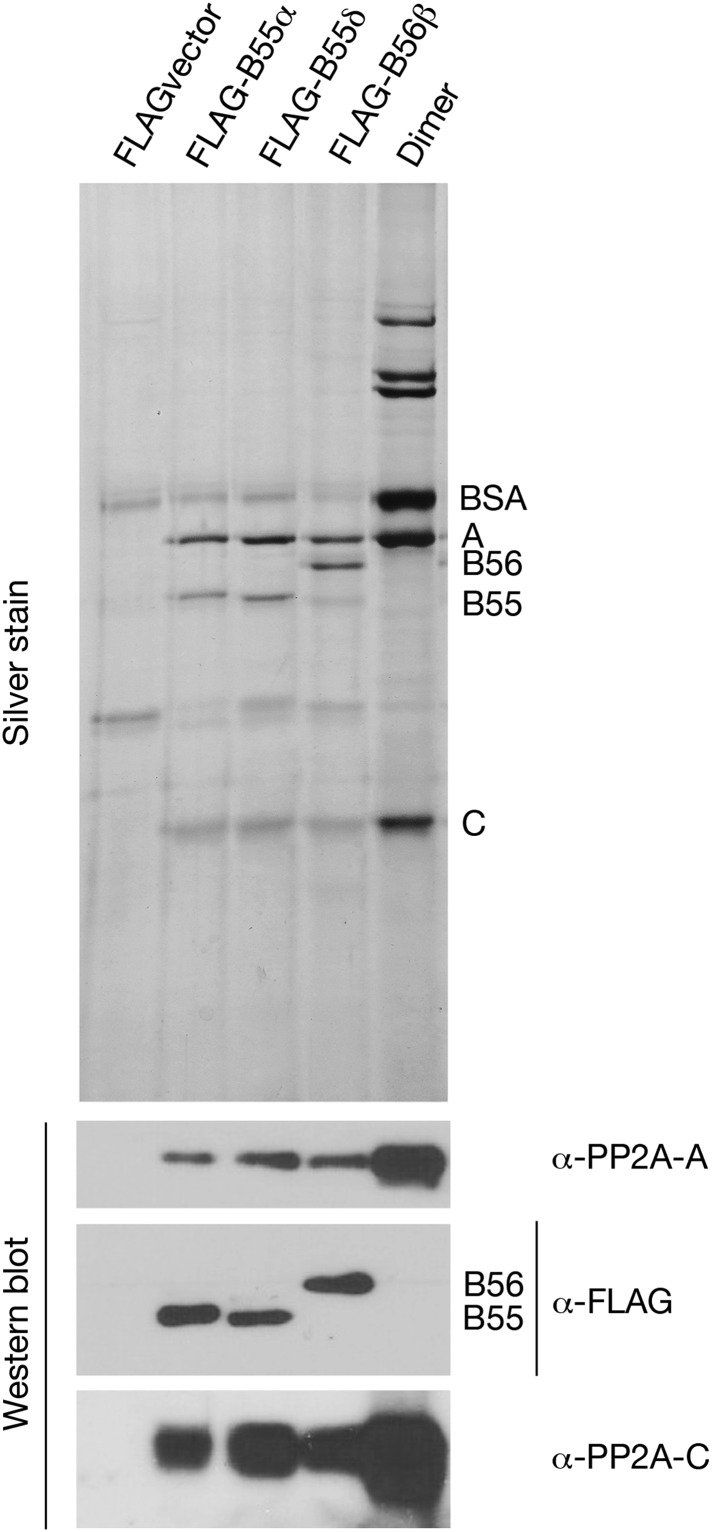
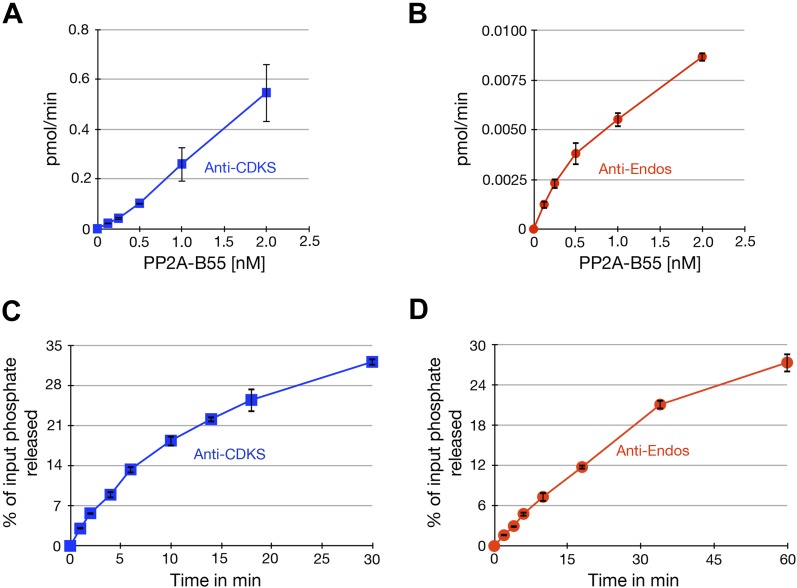
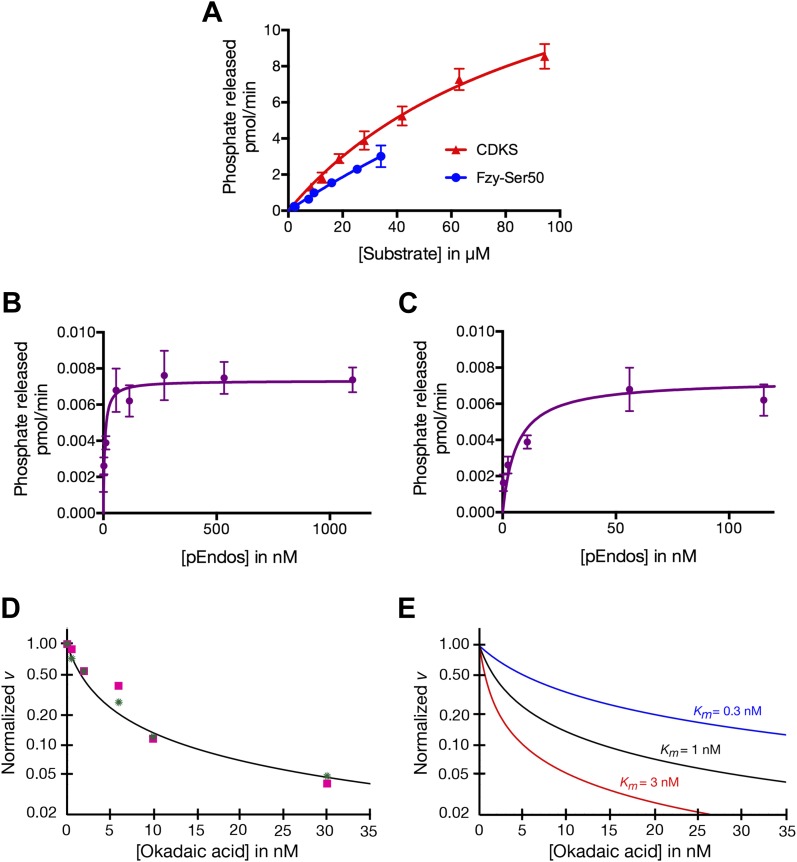
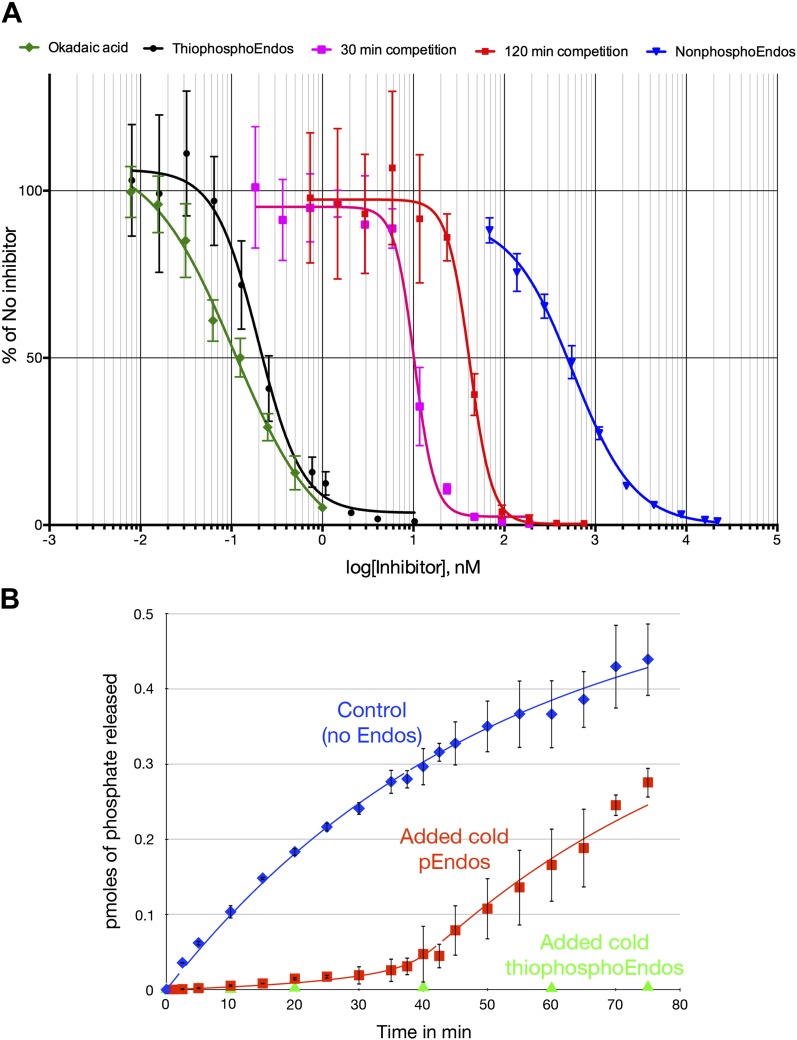
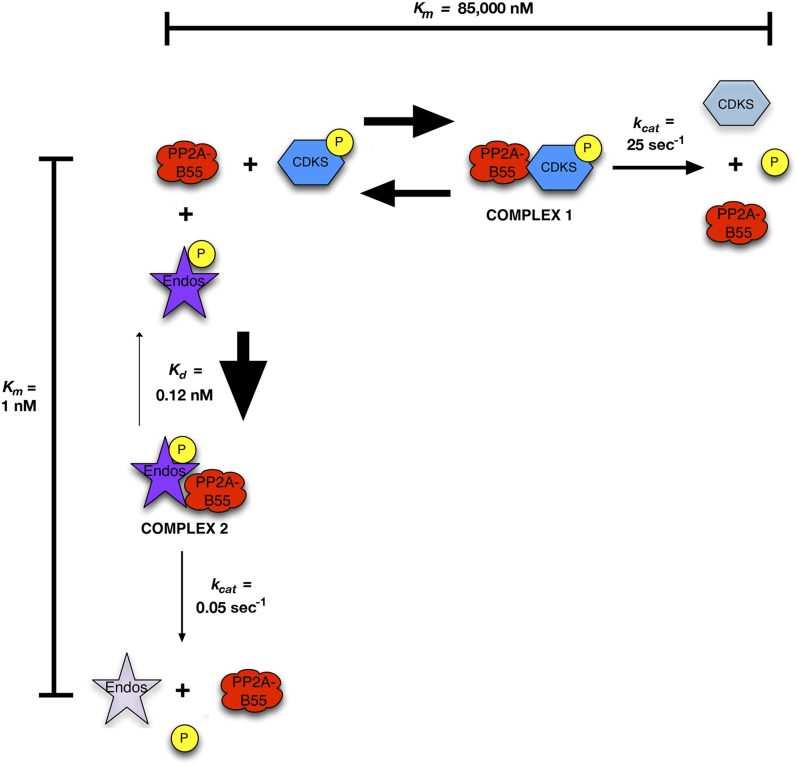
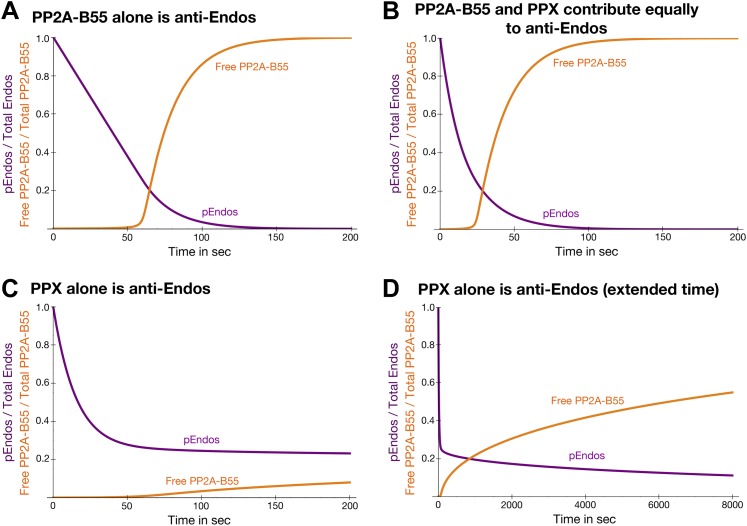
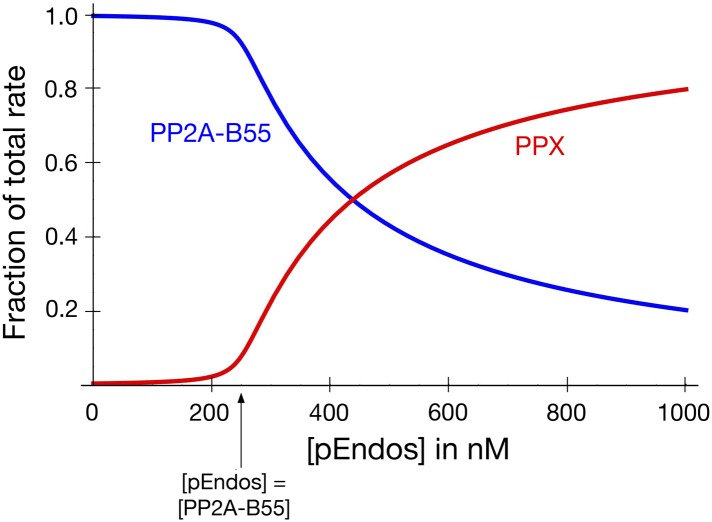
During M phase, Endosulfine (Endos) family proteins are phosphorylated by Greatwall kinase (Gwl), and the resultant pEndos inhibits the phosphatase PP2A-B55, which would otherwise prematurely reverse many CDK-driven phosphorylations. We show here that PP2A-B55 is the enzyme responsible for dephosphorylating pEndos during M phase exit. The kinetic parameters for PP2A-B55's action on pEndos are orders of magnitude lower than those for CDK-phosphorylated substrates, suggesting a simple model for PP2A-B55 regulation that we call inhibition by unfair competition. As the name suggests, during M phase PP2A-B55's attention is diverted to pEndos, which binds much more avidly and is dephosphorylated more slowly than other substrates. When Gwl is inactivated during the M phase-to-interphase transition, the dynamic balance changes: pEndos dephosphorylated by PP2A-B55 cannot be replaced, so the phosphatase can refocus its attention on CDK-phosphorylated substrates. This mechanism explains simultaneously how PP2A-B55 and Gwl together regulate pEndos, and how pEndos controls PP2A-B55. DOI: http://dx.doi.org/10.7554/eLife.01695.001.
在M期,硫丹(Endos)家族蛋白被长城激酶(Gwl)磷酸化,产生的磷酸化硫丹(pEndos)抑制磷酸酶PP2A - B55,否则该磷酸酶会过早逆转许多细胞周期蛋白依赖性激酶(CDK)驱动的磷酸化反应。我们在此表明,PP2A - B55是在M期退出过程中负责使pEndos去磷酸化的酶。PP2A - B55作用于pEndos的动力学参数比其作用于CDK磷酸化底物的动力学参数低几个数量级,这提示了一种我们称为不公平竞争抑制的PP2A - B55调节简单模型。顾名思义,在M期,PP2A - B55的注意力被转移到pEndos上,pEndos比其他底物结合得更紧密且去磷酸化更慢。当Gwl在M期向间期转变过程中失活时,动态平衡发生变化:被PP2A - B55去磷酸化的pEndos无法被替代,因此该磷酸酶能够将注意力重新集中到CDK磷酸化的底物上。这一机制同时解释了PP2A - B55和Gwl如何共同调节pEndos,以及pEndos如何控制PP2A - B55。DOI: http://dx.doi.org/10.7554/eLife.01695.001 。