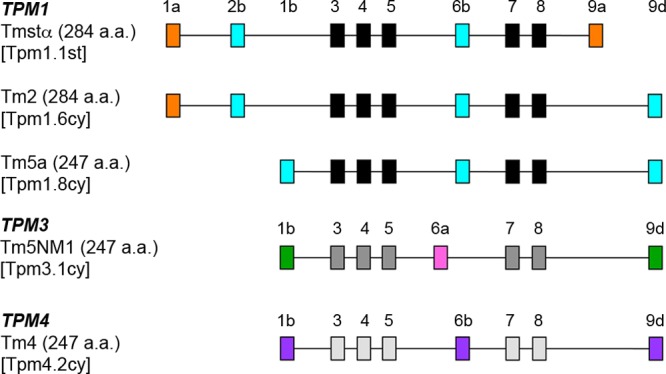
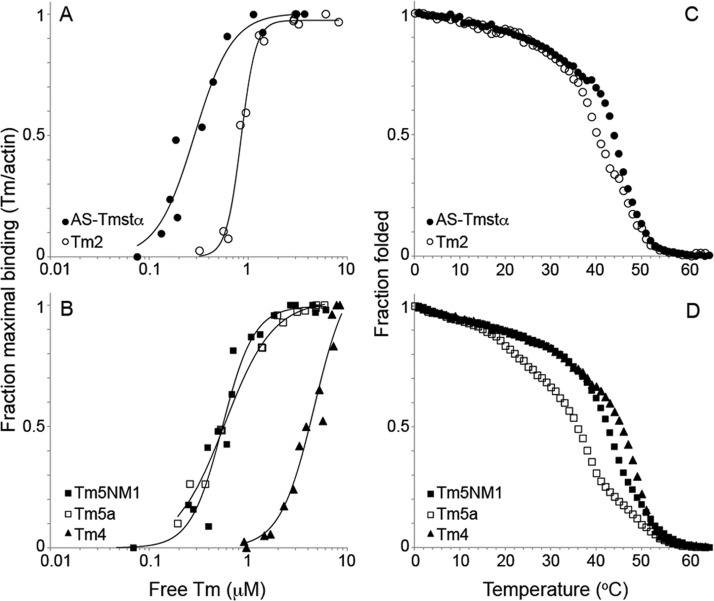
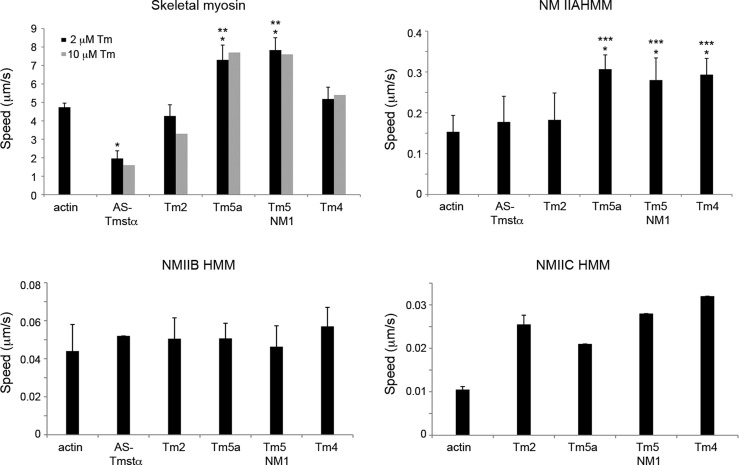
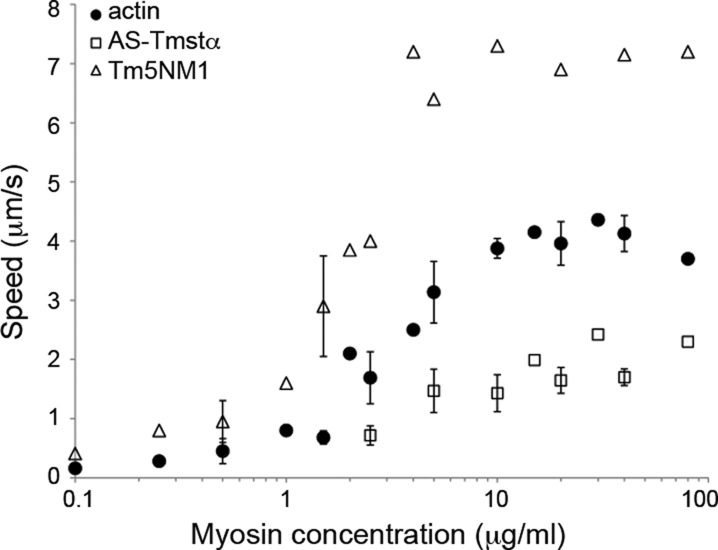
Department of Pathology and Laboratory Medicine, Robert Wood Johnson Medical School, Rutgers University , Piscataway, New Jersey 08854, United States.
Biochemistry. 2014 Jun 24;53(24):4015-24. doi: 10.1021/bi500162z. Epub 2014 Jun 12.
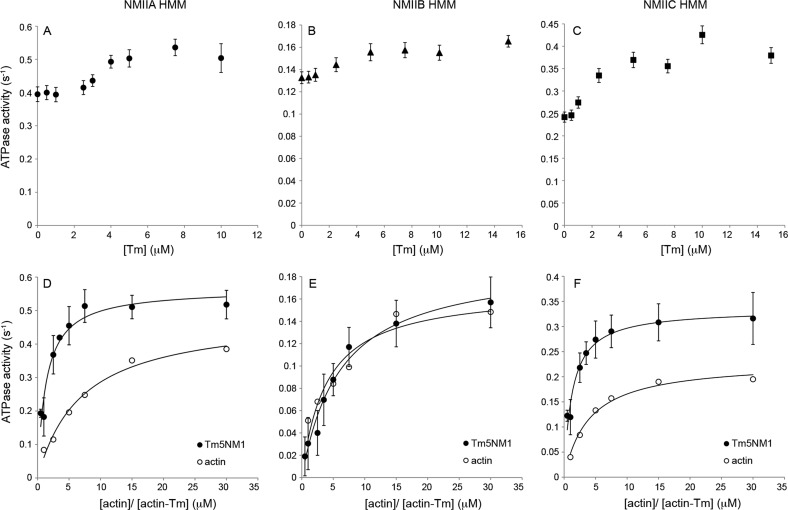
The actin cytoskeleton carries out cellular functions, including division, migration, adhesion, and intracellular transport, that require a variety of actin binding proteins, including myosins. Our focus here is on class II nonmuscle myosin isoforms, NMIIA, NMIIB, and NMIIC, and their regulation by the actin binding protein, tropomyosin. NMII myosins are localized to different populations of stress fibers and the contractile ring, structures involved in force generation required for cell migration, adhesion, and cytokinesis. The stress fibers and contractile ring that contain NMII myosins also contain tropomyosin. Four mammalian genes encode more than 40 tropomyosins. Tropomyosins inhibit or activate actomyosin MgATPase and motility depending on the myosin and tropomyosin isoform. In vivo, tropomyosins play a role in cell migration, adhesion, cytokinesis, and NMII isoform localization in an isoform-specific manner. We postulate that the isoform-specific tropomyosin localization and effect on NMII isoform localization reflect modulation of NMII actomyosin kinetics and motile function. In this study, we compare the ability of different tropomyosin isoforms to support actin filament motility with NMIIA, NMIIB, and NMIIC as well as skeletal muscle myosin. Tropomyosins activated, inhibited, or had no effect on motility depending on the myosin, indicating that the myosin isoform is the primary determinant of the isoform-specific effect of tropomyosin on actomyosin regulation. Activation of motility of nonmuscle tropomyosin-actin filaments by NMII myosin correlates with an increased Vmax of the myosin MgATPase, implying a direct effect on the myosin MgATPase, in contrast to the skeletal tropomyosin-actin filament that has no effect on the Vmax or maximal filament velocity.
肌动蛋白细胞骨架执行细胞功能,包括分裂、迁移、黏附和细胞内运输,这需要多种肌动蛋白结合蛋白,包括肌球蛋白。我们这里的重点是 II 类非肌肉肌球蛋白同工型 NMIIA、NMIIB 和 NMIIC 及其与肌动蛋白结合蛋白原肌球蛋白的调节。NMII 肌球蛋白定位于不同的应力纤维和收缩环种群,这些结构参与细胞迁移、黏附和胞质分裂所需的力生成。含有 NMII 肌球蛋白的应力纤维和收缩环也含有原肌球蛋白。四种哺乳动物基因编码超过 40 种原肌球蛋白。原肌球蛋白根据肌球蛋白和原肌球蛋白同工型抑制或激活肌动球蛋白 MgATP 酶和运动。在体内,原肌球蛋白以同工型特异性方式在细胞迁移、黏附和胞质分裂以及 NMII 同工型定位中发挥作用。我们假设同工型特异性原肌球蛋白定位和对 NMII 同工型定位的影响反映了 NMII 肌球蛋白丝动力学和运动功能的调节。在这项研究中,我们比较了不同原肌球蛋白同工型与 NMIIA、NMIIB 和 NMIIC 以及骨骼肌肌球蛋白一起支持肌动蛋白丝运动的能力。原肌球蛋白根据肌球蛋白激活、抑制或对运动没有影响,表明肌球蛋白同工型是原肌球蛋白对肌球蛋白调节的同工型特异性效应的主要决定因素。非肌肉原肌球蛋白-肌动蛋白丝的 NMII 肌球蛋白的运动激活与肌球蛋白 MgATP 酶的 Vmax 增加相关,这意味着对肌球蛋白 MgATP 酶有直接影响,与骨骼肌原肌球蛋白-肌动蛋白丝相反,它对 Vmax 或最大纤维速度没有影响。