Yang Lei, Brunsfeld John, Scott LuAnn, Wichman Holly
Department of Biological Sciences, University of Idaho, Moscow, Idaho, United States of America; Institute for Bioinformatics and Evolutionary Studies, University of Idaho, Moscow, Idaho, United States of America.
Department of Biological Sciences, University of Idaho, Moscow, Idaho, United States of America.
PLoS Genet. 2014 Jun 26;10(6):e1004395. doi: 10.1371/journal.pgen.1004395. eCollection 2014 Jun.
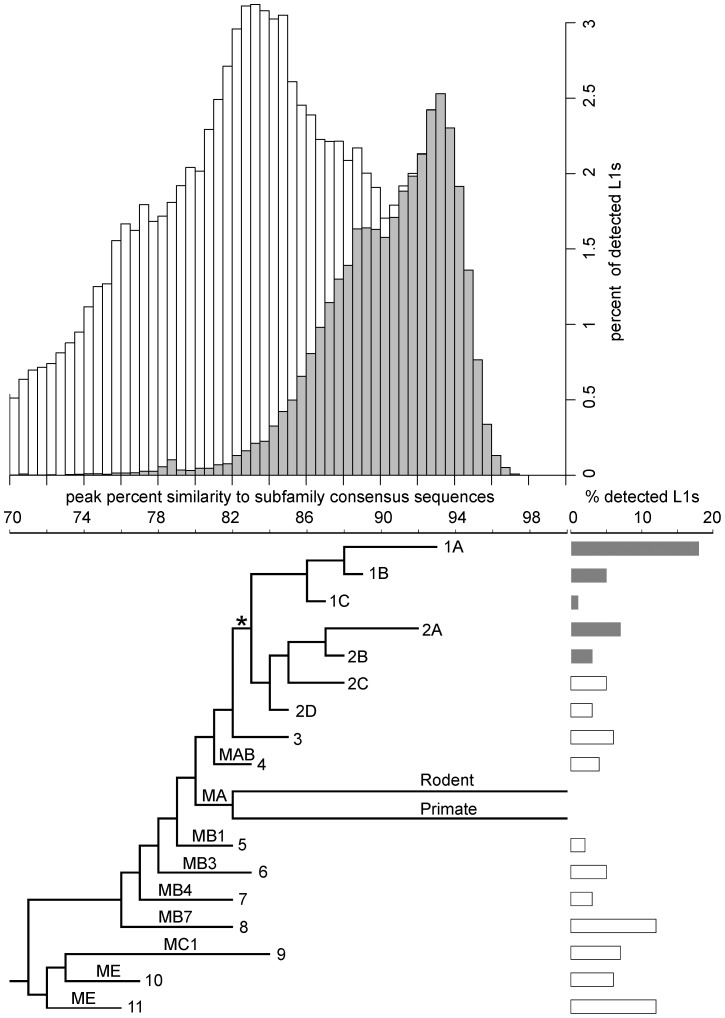
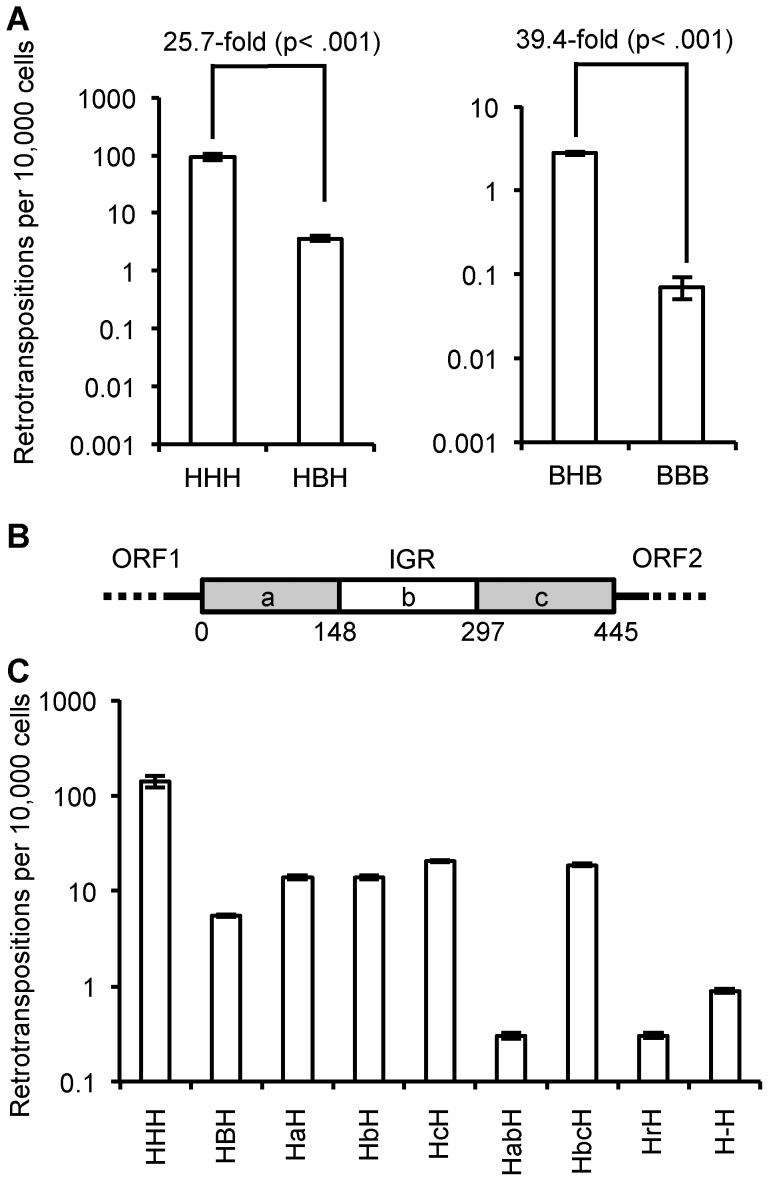
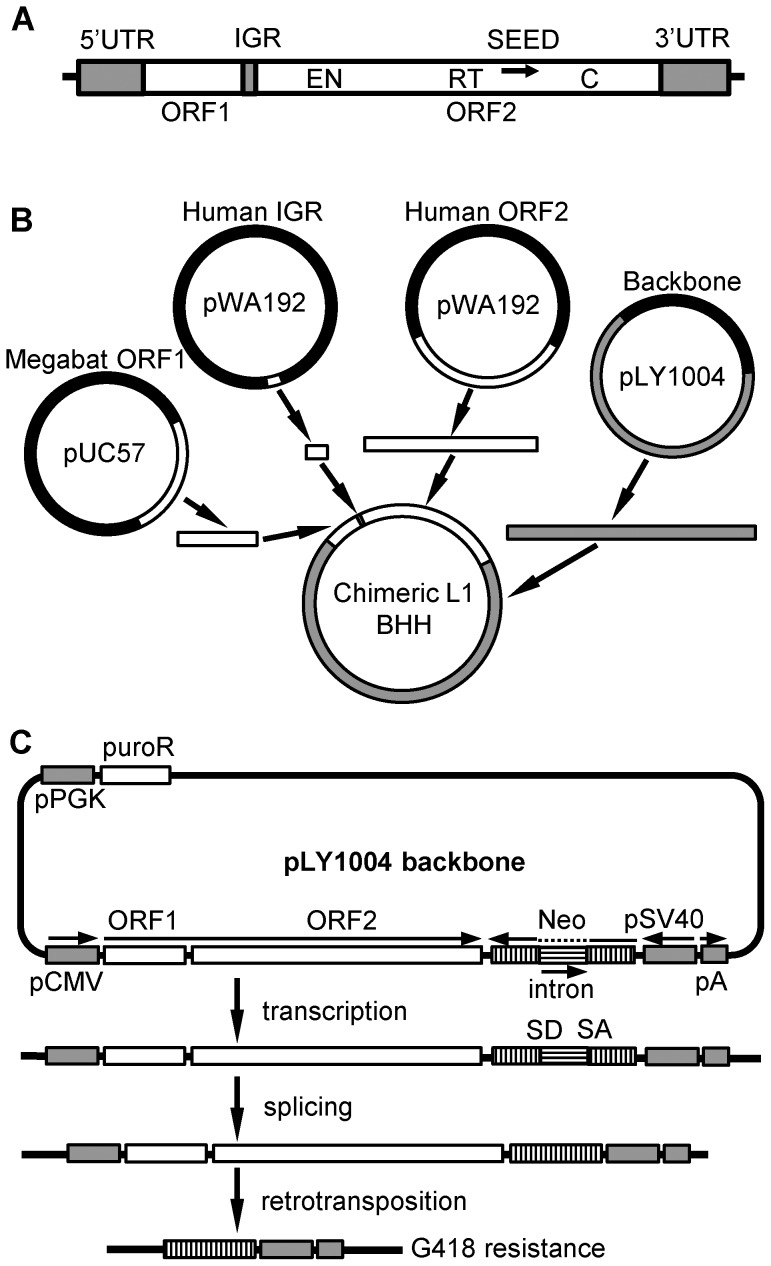
Although L1 sequences are present in the genomes of all placental mammals and marsupials examined to date, their activity was lost in the megabat family, Pteropodidae, ∼24 million years ago. To examine the characteristics of L1s prior to their extinction, we analyzed the evolutionary history of L1s in the genome of a megabat, Pteropus vampyrus, and found a pattern of periodic L1 expansion and quiescence. In contrast to the well-characterized L1s in human and mouse, megabat genomes have accommodated two or more simultaneously active L1 families throughout their evolutionary history, and major peaks of L1 deposition into the genome always involved multiple families. We compared the consensus sequences of the two major megabat L1 families at the time of their extinction to consensus L1s of a variety of mammalian species. Megabat L1s are comparable to the other mammalian L1s in terms of adenosine content and conserved amino acids in the open reading frames (ORFs). However, the intergenic region (IGR) of the reconstructed element from the more active family is dramatically longer than the IGR of well-characterized human and mouse L1s. We synthesized the reconstructed element from this L1 family and tested the ability of its components to support retrotransposition in a tissue culture assay. Both ORFs are capable of supporting retrotransposition, while the IGR is inhibitory to retrotransposition, especially when combined with either of the reconstructed ORFs. We dissected the inhibitory effect of the IGR by testing truncated and shuffled versions and found that length is a key factor, but not the only one affecting inhibition of retrotransposition. Although the IGR is inhibitory to retrotransposition, this inhibition does not account for the extinction of L1s in megabats. Overall, the evolution of the L1 sequence or the quiescence of L1 is unlikely the reason of L1 extinction.
尽管迄今为止在所有已检测的胎盘哺乳动物和有袋动物的基因组中都存在L1序列,但在约2400万年前,它们的活性在大蝙蝠科(狐蝠科)中丧失。为了研究L1在灭绝之前的特征,我们分析了一种大蝙蝠(马来大狐蝠)基因组中L1的进化历史,发现了L1周期性扩张和静止的模式。与人类和小鼠中特征明确的L1不同,大蝙蝠基因组在其整个进化历史中容纳了两个或更多同时活跃的L1家族,并且L1整合到基因组中的主要峰值总是涉及多个家族。我们将两种主要的大蝙蝠L1家族在灭绝时的共有序列与多种哺乳动物物种的共有L1序列进行了比较。大蝙蝠L1在腺苷含量和开放阅读框(ORF)中的保守氨基酸方面与其他哺乳动物L1相当。然而,来自更活跃家族的重建元件的基因间区域(IGR)比特征明确的人类和小鼠L1的IGR长得多。我们合成了来自这个L1家族的重建元件,并在组织培养试验中测试了其组件支持逆转录转座的能力。两个ORF都能够支持逆转录转座,而IGR对逆转录转座具有抑制作用,尤其是与任何一个重建的ORF结合时。我们通过测试截短和改组版本来剖析IGR的抑制作用,发现长度是一个关键因素,但不是影响逆转录转座抑制的唯一因素。尽管IGR对逆转录转座具有抑制作用,但这种抑制并不能解释大蝙蝠中L1的灭绝。总体而言,L1序列的进化或L1的静止不太可能是L1灭绝的原因。