Hodges Jennifer L, Vilchez Samuel Martin, Asmussen Hannelore, Whitmore Leanna A, Horwitz Alan Rick
Department of Cell Biology, University of Virginia School of Medicine, Charlottesville, Virginia, United States of America.
PLoS One. 2014 Jul 9;9(7):e101770. doi: 10.1371/journal.pone.0101770. eCollection 2014.
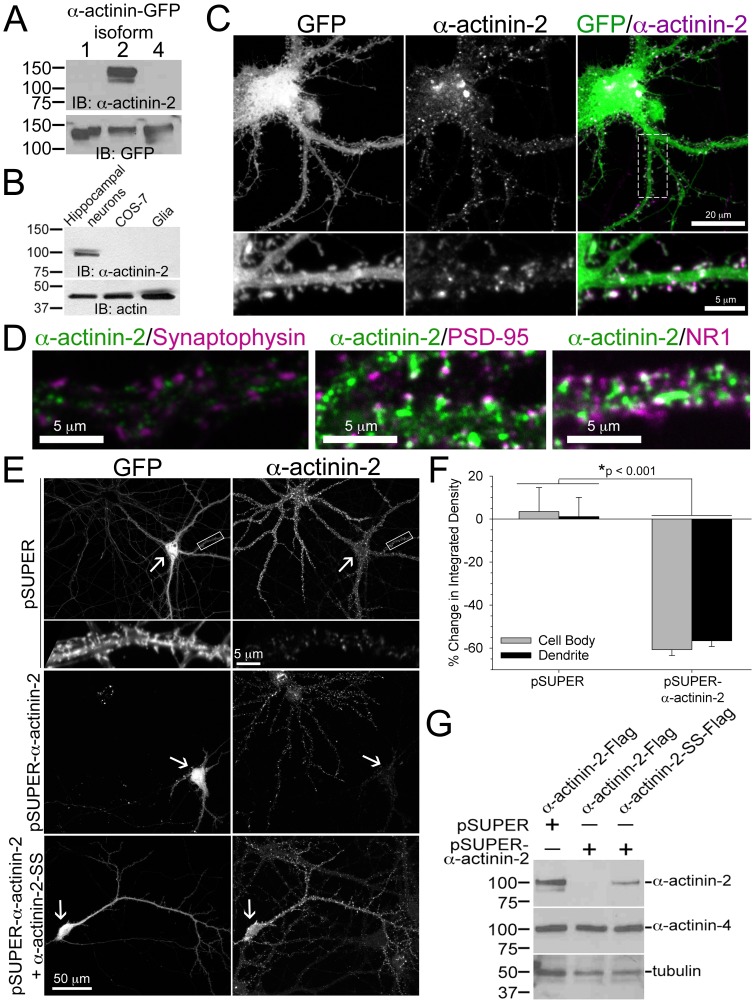
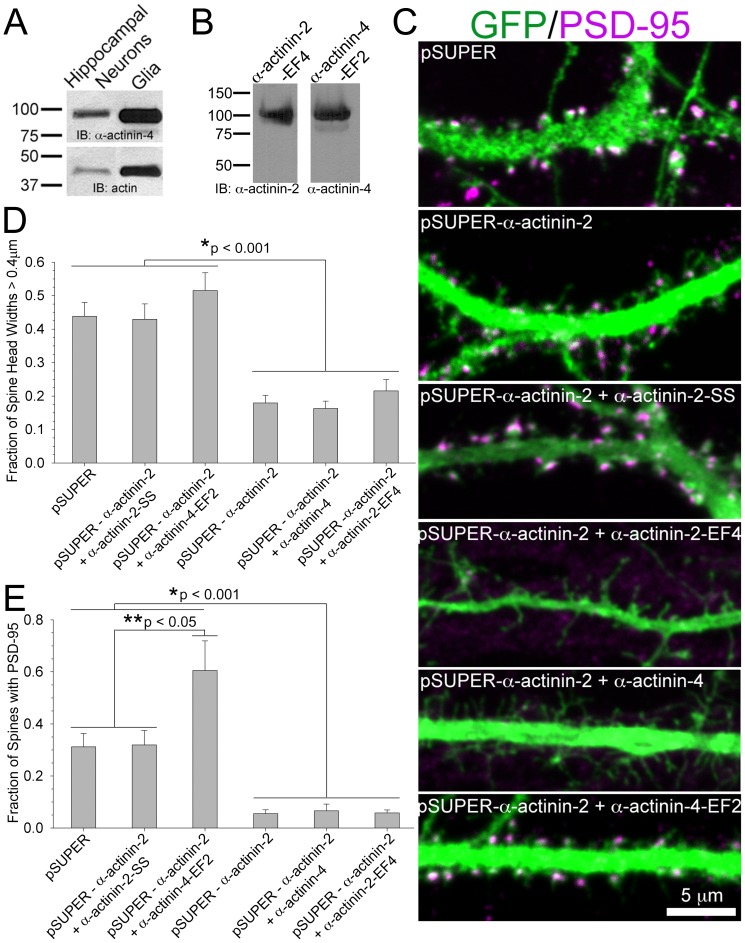
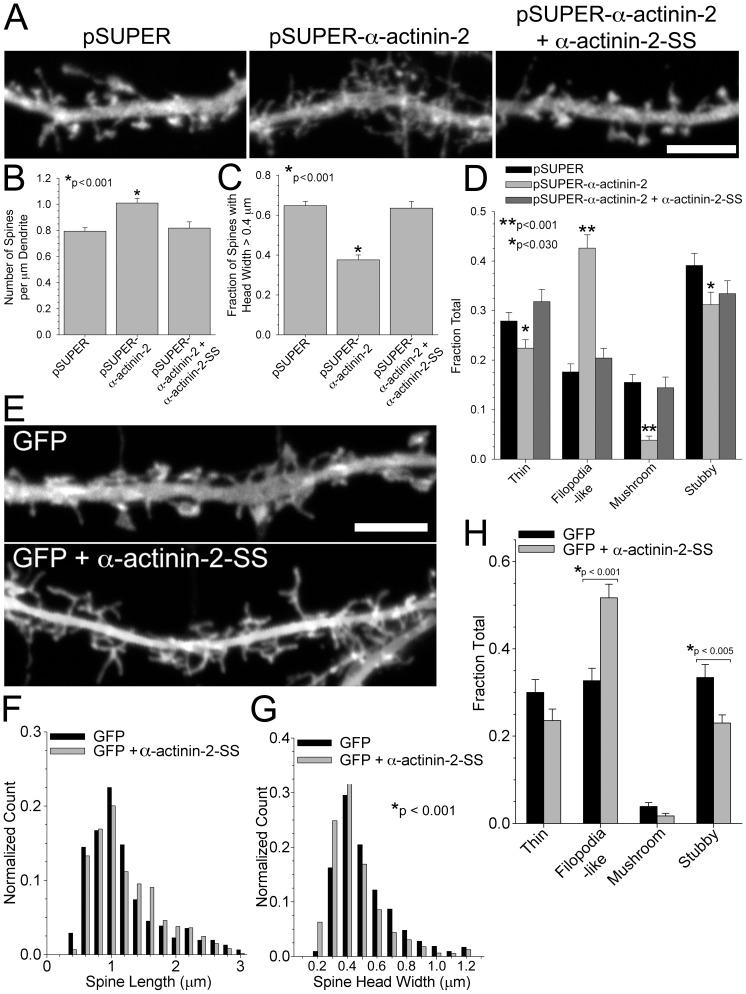
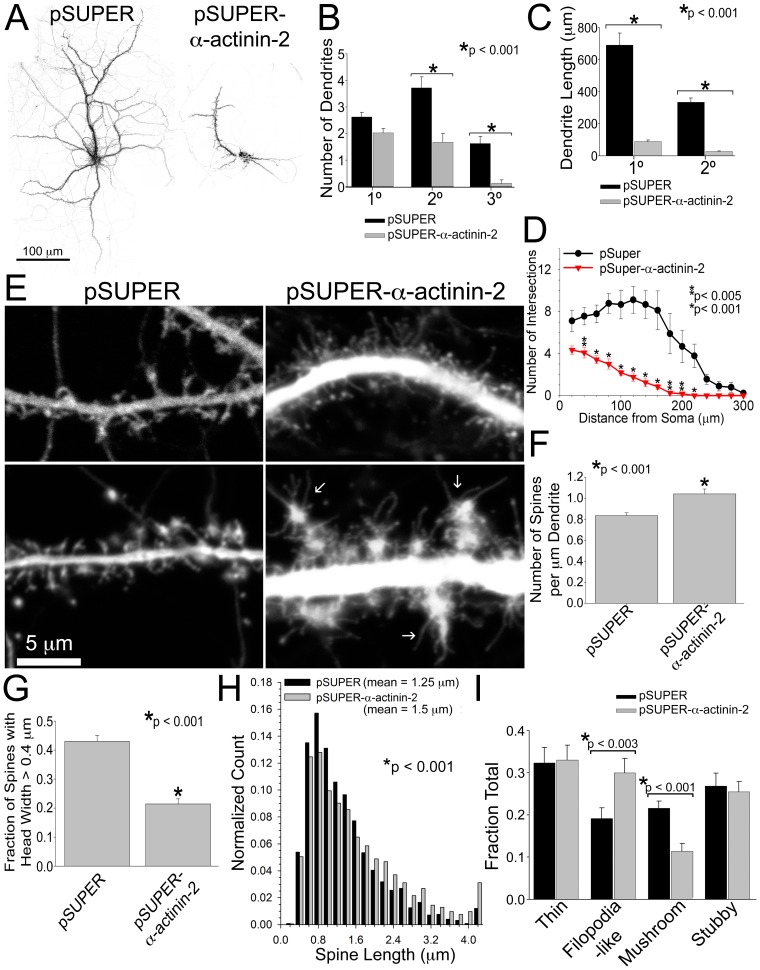
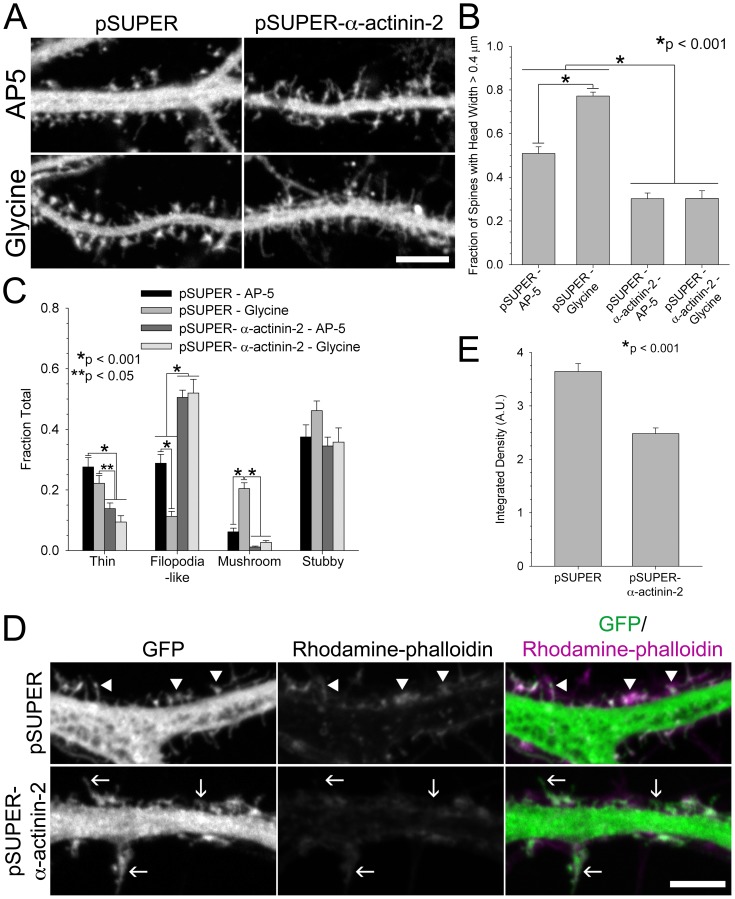
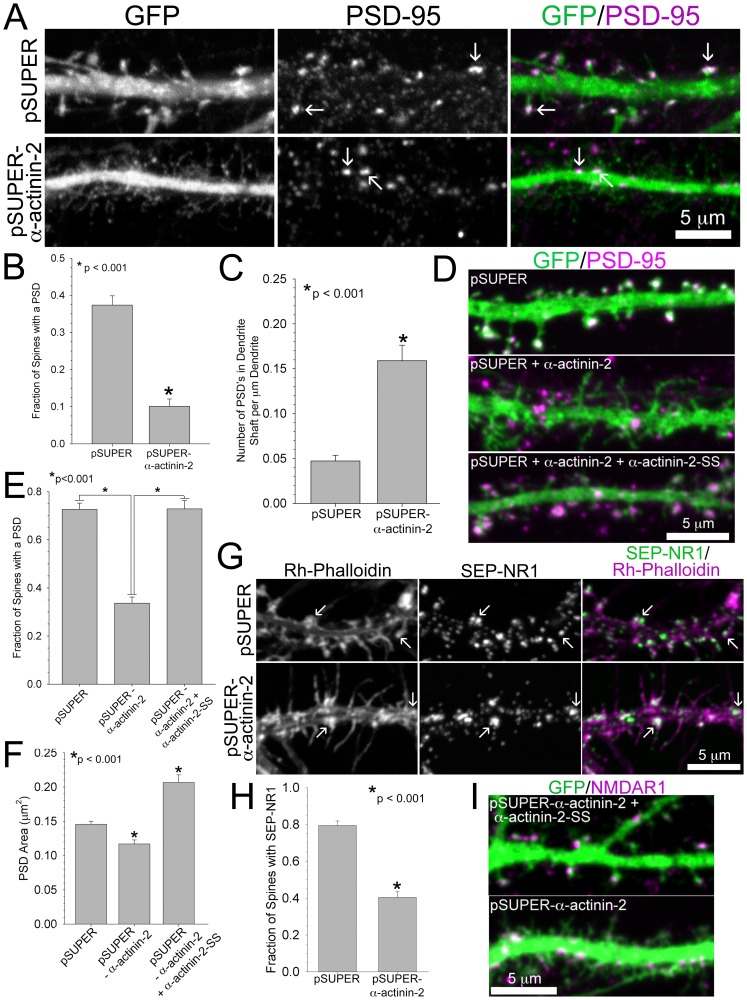
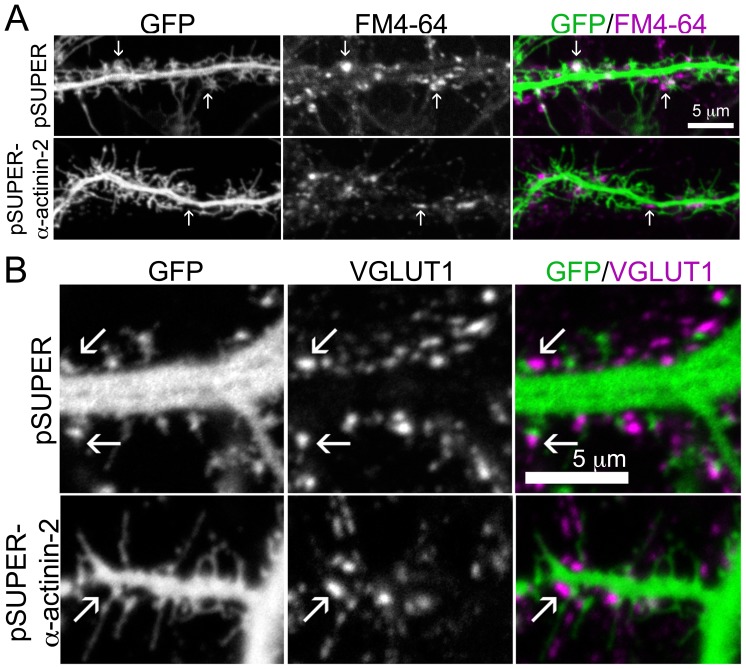
Dendritic spines are micron-sized protrusions that constitute the primary post-synaptic sites of excitatory neurotransmission in the brain. Spines mature from a filopodia-like protrusion into a mushroom-shaped morphology with a post-synaptic density (PSD) at its tip. Modulation of the actin cytoskeleton drives these morphological changes as well as the spine dynamics that underlie learning and memory. Several PSD molecules respond to glutamate receptor activation and relay signals to the underlying actin cytoskeleton to regulate the structural changes in spine and PSD morphology. α-Actinin-2 is an actin filament cross-linker, which localizes to dendritic spines, enriched within the post-synaptic density, and implicated in actin organization. We show that loss of α-actinin-2 in rat hippocampal neurons creates an increased density of immature, filopodia-like protrusions that fail to mature into a mushroom-shaped spine during development. α-Actinin-2 knockdown also prevents the recruitment and stabilization of the PSD in the spine, resulting in failure of synapse formation, and an inability to structurally respond to chemical stimulation of the N-methyl-D-aspartate (NMDA)-type glutamate receptor. The Ca2+-insensitive EF-hand motif in α-actinin-2 is necessary for the molecule's function in regulating spine morphology and PSD assembly, since exchanging it for the similar but Ca2+-sensitive domain from α-actinin-4, another α-actinin isoform, inhibits its function. Furthermore, when the Ca2+-insensitive domain from α-actinin-2 is inserted into α-actinin-4 and expressed in neurons, it creates mature spines. These observations support a model whereby α-actinin-2, partially through its Ca2+-insensitive EF-hand motif, nucleates PSD formation via F-actin organization and modulates spine maturation to mediate synaptogenesis.
树突棘是微米大小的突起,构成大脑中兴奋性神经传递的主要突触后位点。树突棘从丝状伪足样突起发育成熟为蘑菇状形态,其顶端有一个突触后致密区(PSD)。肌动蛋白细胞骨架的调节驱动这些形态变化以及作为学习和记忆基础的树突棘动态变化。几种PSD分子对谷氨酸受体激活作出反应,并将信号传递到下层的肌动蛋白细胞骨架,以调节树突棘和PSD形态的结构变化。α-辅肌动蛋白-2是一种肌动蛋白丝交联蛋白,定位于树突棘,在突触后致密区富集,并与肌动蛋白组织有关。我们发现,大鼠海马神经元中α-辅肌动蛋白-2的缺失导致未成熟的丝状伪足样突起密度增加,这些突起在发育过程中无法成熟为蘑菇状树突棘。敲低α-辅肌动蛋白-2还会阻止PSD在树突棘中的募集和稳定,导致突触形成失败,并且无法对N-甲基-D-天冬氨酸(NMDA)型谷氨酸受体的化学刺激作出结构反应。α-辅肌动蛋白-2中对Ca2+不敏感的EF手基序对于该分子在调节树突棘形态和PSD组装中的功能是必需的,因为将其替换为另一种α-辅肌动蛋白异构体α-辅肌动蛋白-4中相似但对Ca2+敏感的结构域会抑制其功能。此外,当将α-辅肌动蛋白-2中对Ca2+不敏感的结构域插入α-辅肌动蛋白-4并在神经元中表达时,会产生成熟的树突棘。这些观察结果支持了一种模型,即α-辅肌动蛋白-2部分通过其对Ca2+不敏感的EF手基序,通过F-肌动蛋白组织促进PSD形成并调节树突棘成熟以介导突触形成。