Tark-Dame Mariliis, Jerabek Hansjoerg, Manders Erik M M, van der Wateren Ingrid M, Heermann Dieter W, van Driel Roel
Swammerdam Institute for Life Sciences, University of Amsterdam, Amsterdam, The Netherlands.
Institute for Theoretical Physics, Heidelberg University, Heidelberg, Germany.
PLoS Comput Biol. 2014 Oct 9;10(10):e1003877. doi: 10.1371/journal.pcbi.1003877. eCollection 2014 Oct.
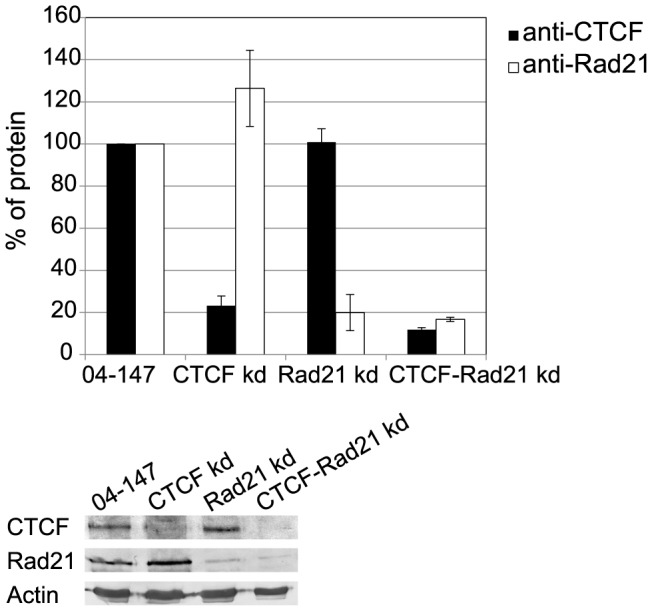
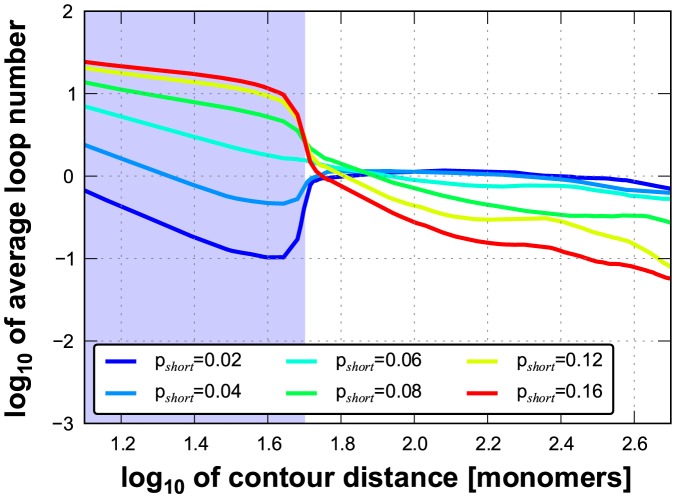
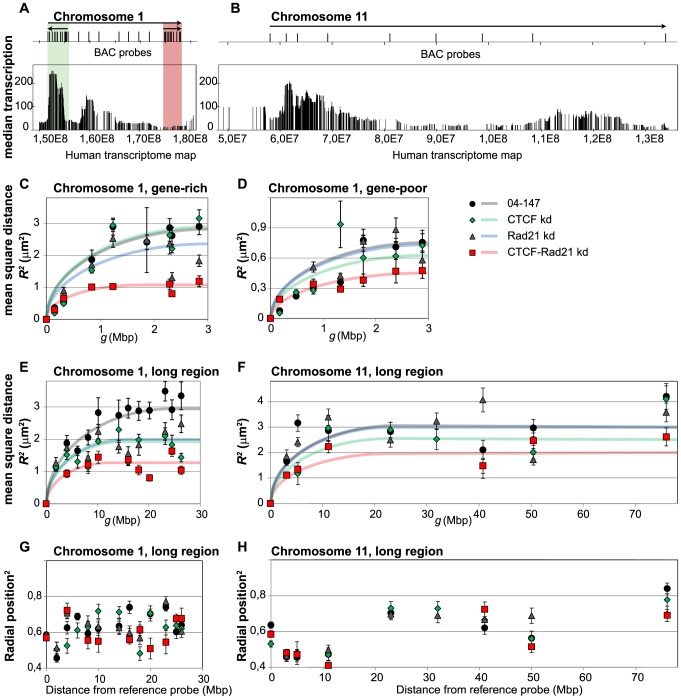
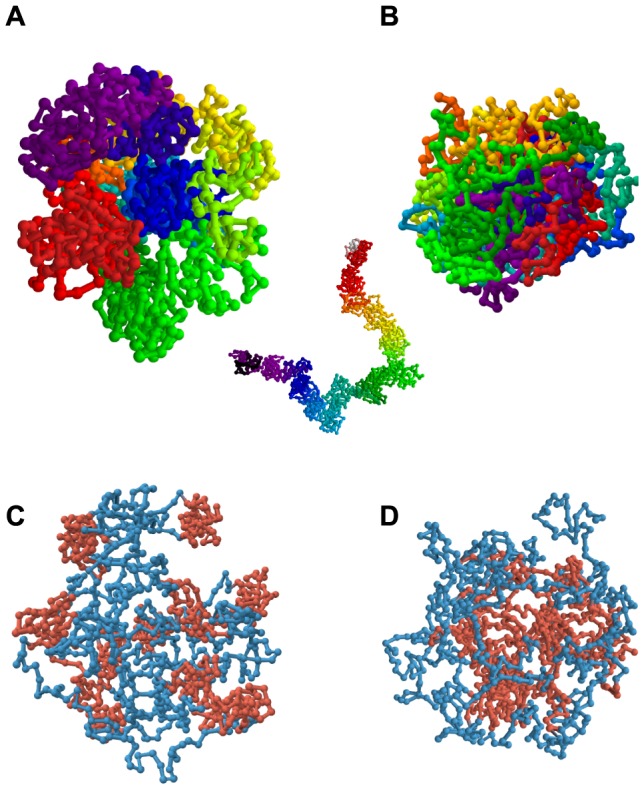
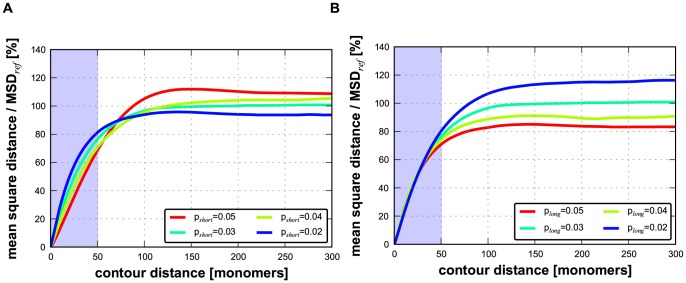
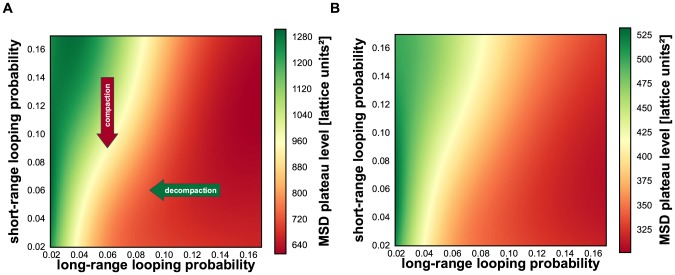
Folding of the chromosomal fibre in interphase nuclei is an important element in the regulation of gene expression. For instance, physical contacts between promoters and enhancers are a key element in cell-type-specific transcription. We know remarkably little about the principles that control chromosome folding. Here we explore the view that intrachromosomal interactions, forming a complex pattern of loops, are a key element in chromosome folding. CTCF and cohesin are two abundant looping proteins of interphase chromosomes of higher eukaryotes. To investigate the role of looping in large-scale (supra Mb) folding of human chromosomes, we knocked down the gene that codes for CTCF and the one coding for Rad21, an essential subunit of cohesin. We measured the effect on chromosome folding using systematic 3D fluorescent in situ hybridization (FISH). Results show that chromatin becomes more compact after reducing the concentration of these two looping proteins. The molecular basis for this counter-intuitive behaviour is explored by polymer modelling usingy the Dynamic Loop model (Bohn M, Heermann DW (2010) Diffusion-driven looping provides a consistent framework for chromatin organization. PLoS ONE 5: e12218.). We show that compaction can be explained by selectively decreasing the number of short-range loops, leaving long-range looping unchanged. In support of this model prediction it has recently been shown by others that CTCF and cohesin indeed are responsible primarily for short-range looping. Our results suggest that the local and the overall changes in of chromosome structure are controlled by a delicate balance between short-range and long-range loops, allowing easy switching between, for instance, open and more compact chromatin states.
间期细胞核中染色体纤维的折叠是基因表达调控的一个重要因素。例如,启动子与增强子之间的物理接触是细胞类型特异性转录的关键因素。然而,我们对控制染色体折叠的原理知之甚少。在这里,我们探讨了一种观点,即形成复杂环模式的染色体内相互作用是染色体折叠的关键因素。CTCF和黏连蛋白是高等真核生物间期染色体中两种丰富的环形成蛋白。为了研究环形成在人类染色体大规模(超百万碱基对)折叠中的作用,我们敲低了编码CTCF的基因以及编码黏连蛋白的一个必需亚基Rad21的基因。我们使用系统的三维荧光原位杂交(FISH)来测量对染色体折叠的影响。结果表明,降低这两种环形成蛋白的浓度后,染色质变得更加致密。我们通过使用动态环模型(Bohn M, Heermann DW (2010) Diffusion-driven looping provides a consistent framework for chromatin organization. PLoS ONE 5: e12218.)的聚合物建模来探索这种与直觉相反行为的分子基础。我们表明,染色质致密化可以通过选择性减少短程环的数量来解释,而长程环不受影响。最近其他人的研究表明CTCF和黏连蛋白确实主要负责短程环的形成,这支持了我们的模型预测。我们的结果表明,染色体结构的局部和整体变化是由短程环和长程环之间的微妙平衡控制的,这使得例如开放和更致密的染色质状态之间能够轻松切换。