Koka Kanthaiah, Tollin Daniel J
Department of Physiology and Biophysics, University of Colorado School of Medicine Aurora, CO, USA.
Front Neural Circuits. 2014 Dec 16;8:144. doi: 10.3389/fncir.2014.00144. eCollection 2014.
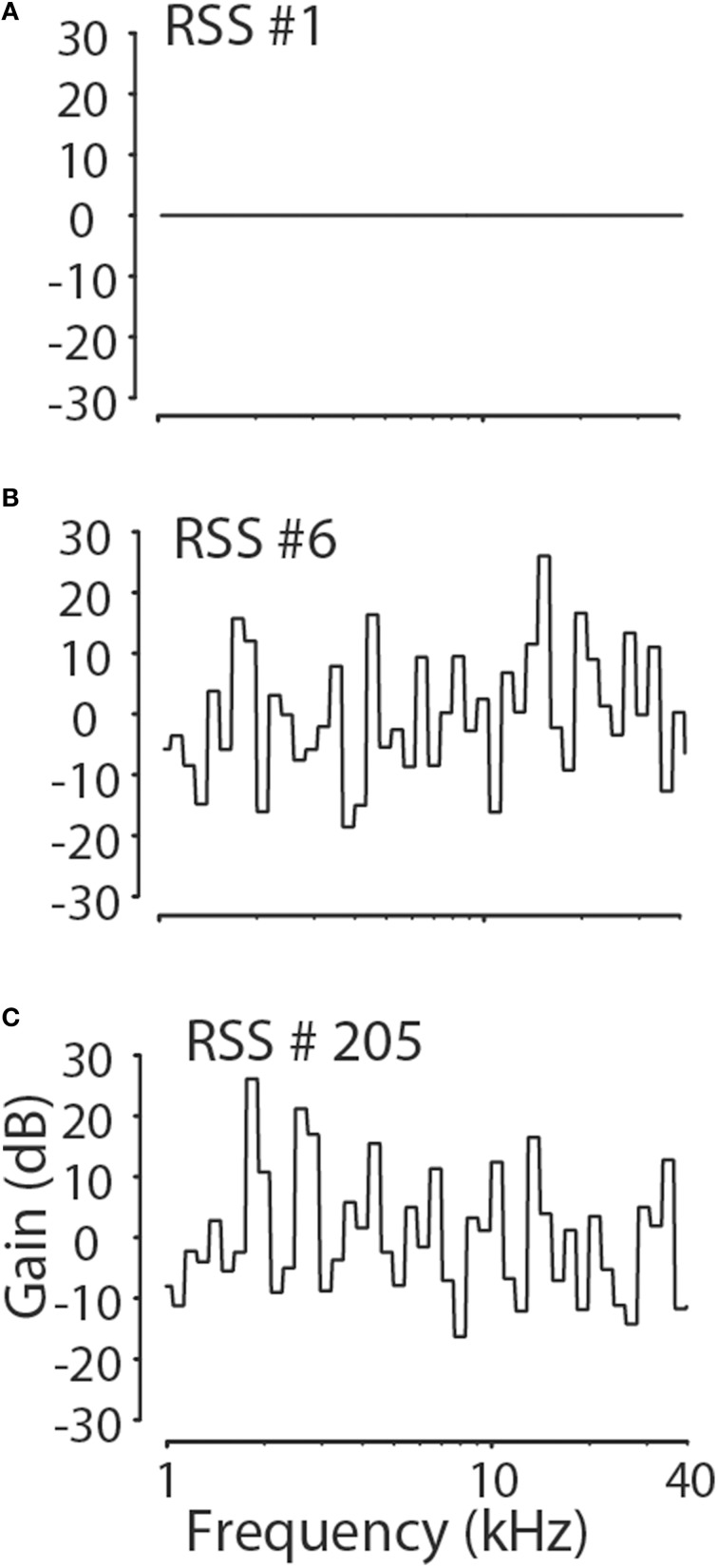
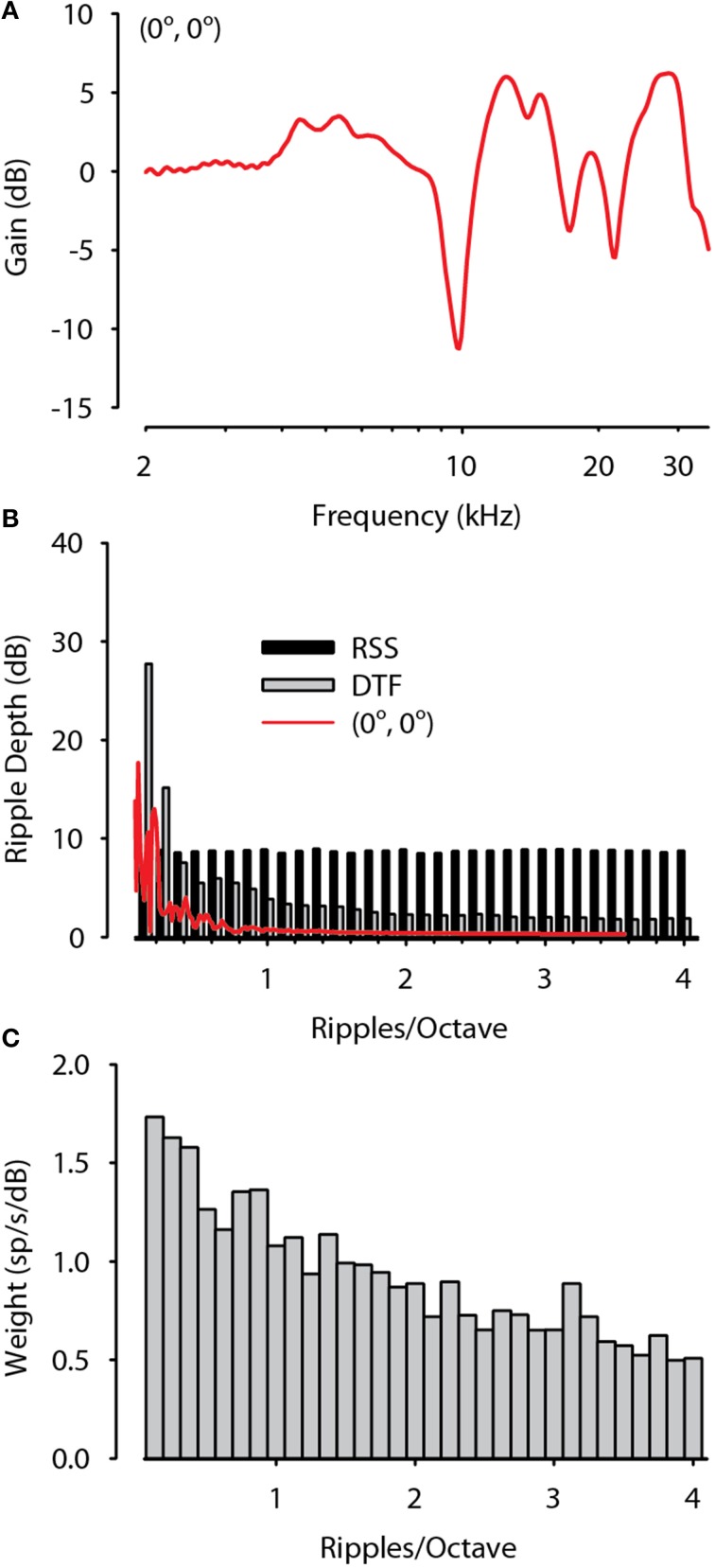
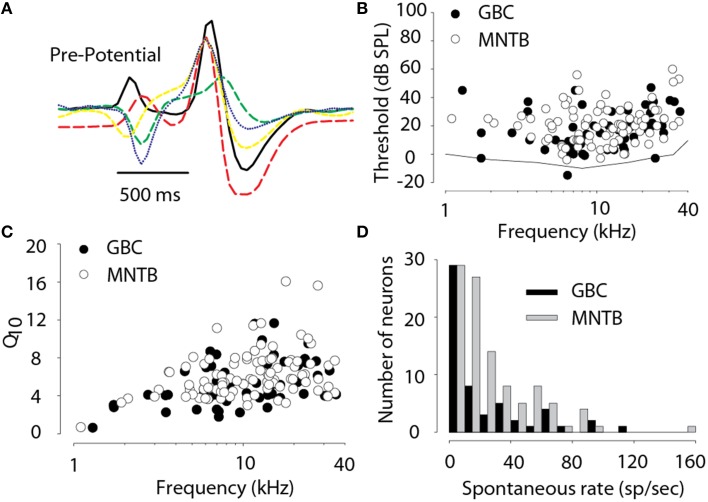
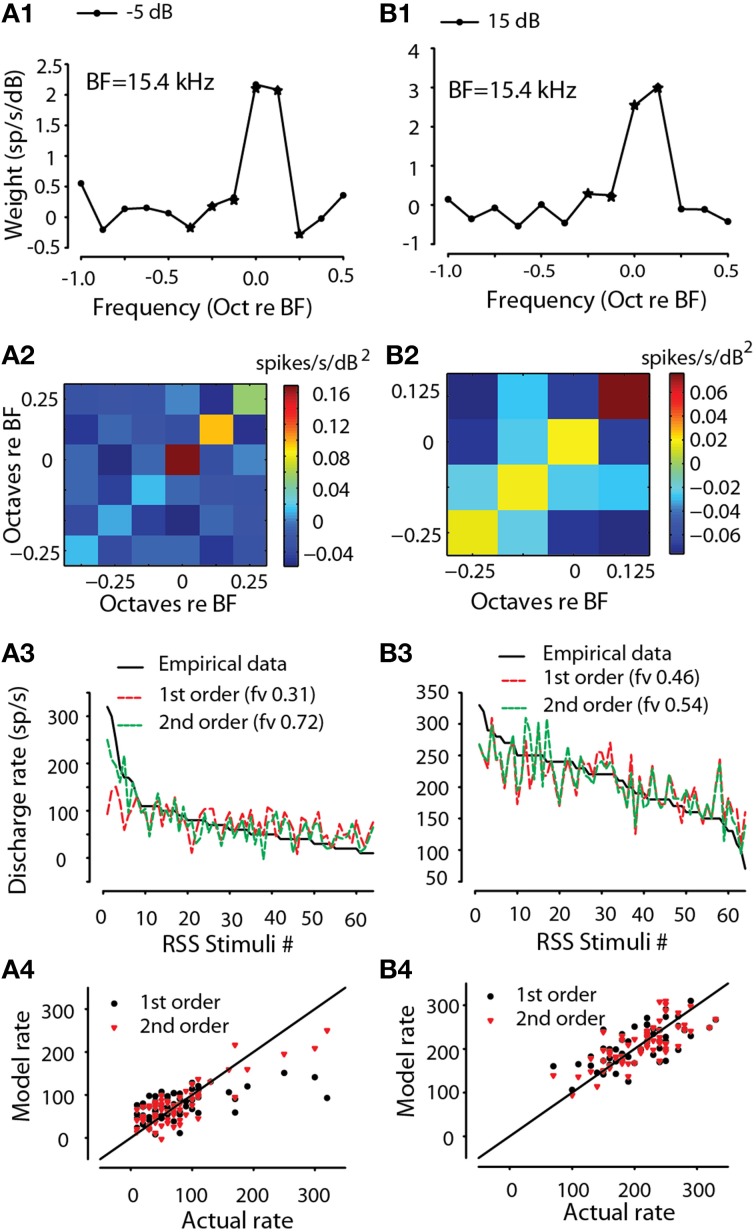
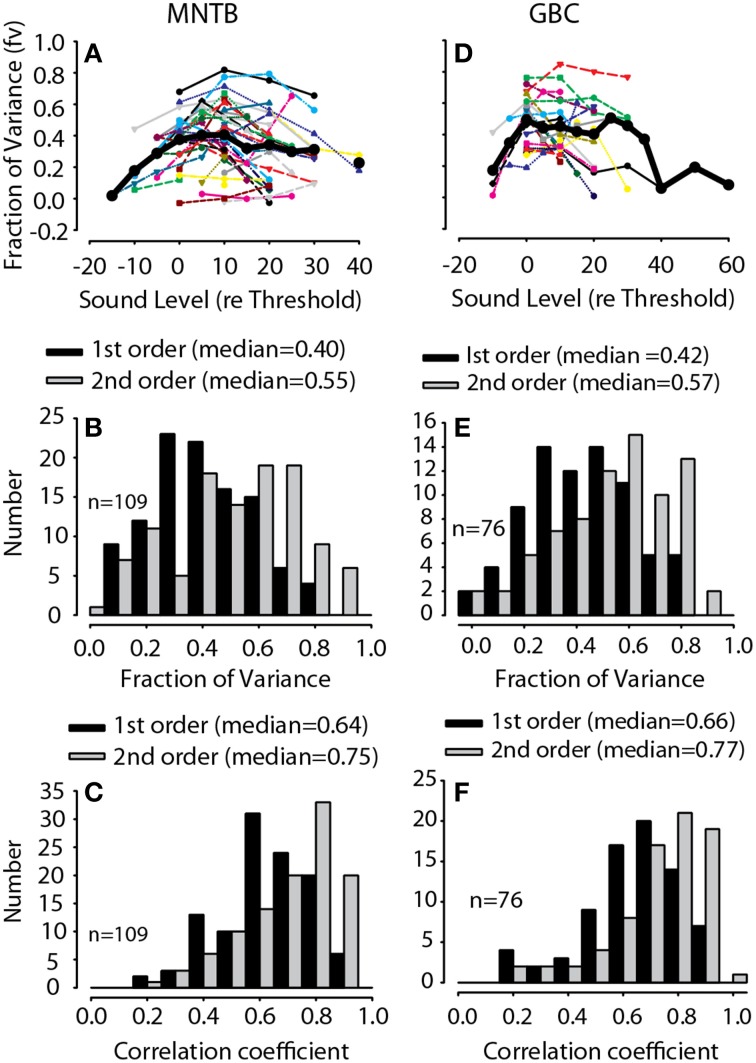
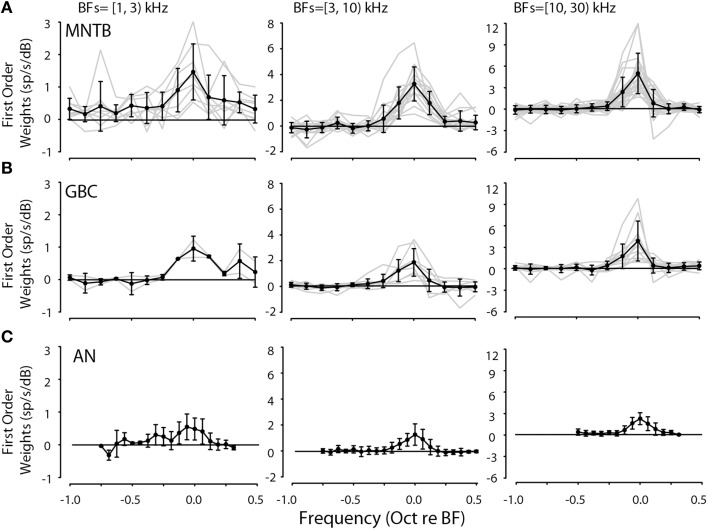
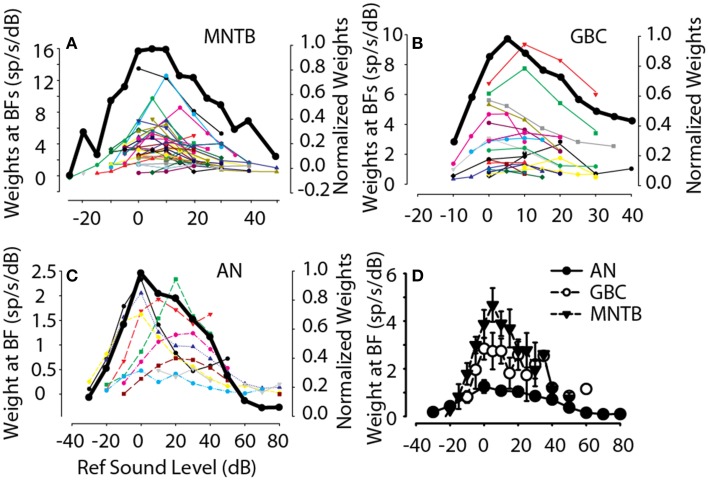
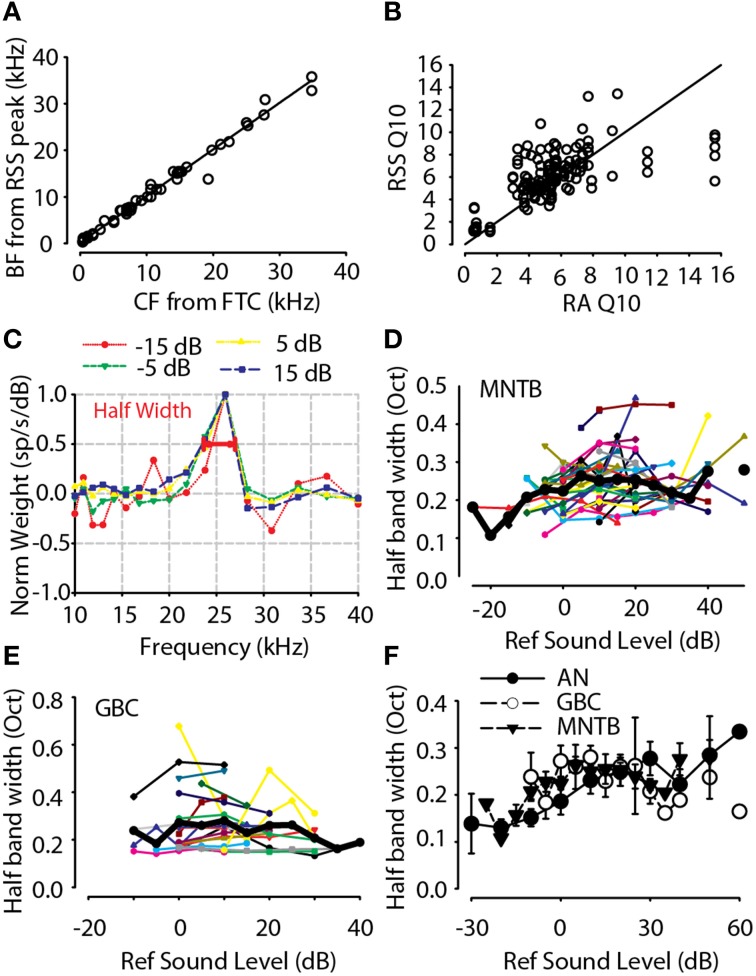
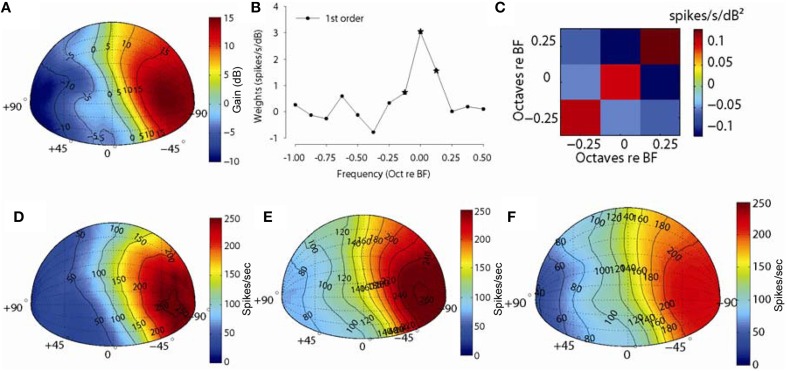
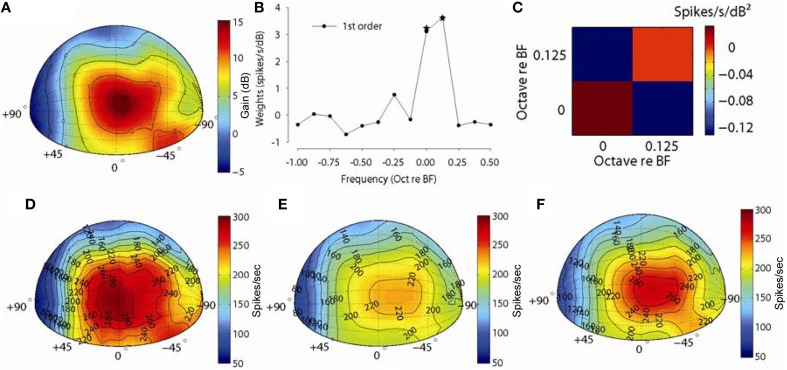
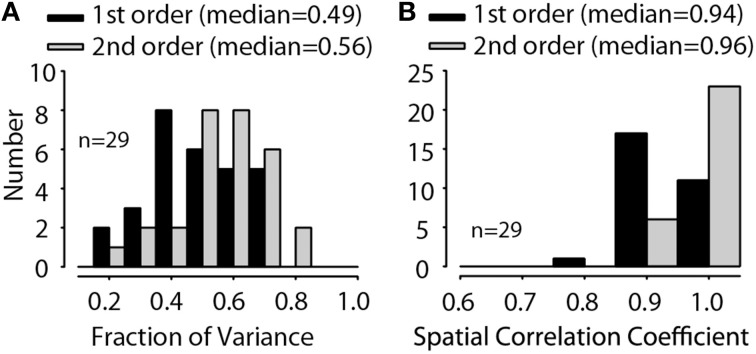
The interaural level difference (ILD) cue to sound location is first encoded in the lateral superior olive (LSO). ILD sensitivity results because the LSO receives excitatory input from the ipsilateral cochlear nucleus and inhibitory input indirectly from the contralateral cochlear nucleus via glycinergic neurons of the ipsilateral medial nucleus of the trapezoid body (MNTB). It is hypothesized that in order for LSO neurons to encode ILDs, the sound spectra at both ears must be accurately encoded via spike rate by their afferents. This spectral-coding hypothesis has not been directly tested in MNTB, likely because MNTB neurons have been mostly described and studied recently in regards to their abilities to encode temporal aspects of sounds, not spectral. Here, we test the hypothesis that MNTB neurons and their inputs from the cochlear nucleus and auditory nerve code sound spectra via discharge rate. The Random Spectral Shape (RSS) method was used to estimate how the levels of 100-ms duration spectrally stationary stimuli were weighted, both linearly and non-linearly, across a wide band of frequencies. In general, MNTB neurons, and their globular bushy cell inputs, were found to be well-modeled by a linear weighting of spectra demonstrating that the pathways through the MNTB can accurately encode sound spectra including those resulting from the acoustical cues to sound location provided by head-related directional transfer functions (DTFs). Together with the anatomical and biophysical specializations for timing in the MNTB-LSO complex, these mechanisms may allow ILDs to be computed for complex stimuli with rapid spectrotemporally-modulated envelopes such as speech and animal vocalizations and moving sound sources.
声音定位的双耳声级差(ILD)线索首先在外侧上橄榄核(LSO)中进行编码。产生ILD敏感性的原因是,LSO接收来自同侧耳蜗核的兴奋性输入以及经由同侧斜方体内侧核(MNTB)的甘氨酸能神经元间接来自对侧耳蜗核的抑制性输入。据推测,为了使LSO神经元对ILD进行编码,双耳的声谱必须由其传入神经通过发放率进行精确编码。这一频谱编码假说尚未在MNTB中得到直接验证,可能是因为MNTB神经元最近大多是在其编码声音时间特征而非频谱特征的能力方面被描述和研究。在这里,我们验证这一假说,即MNTB神经元及其来自耳蜗核和听神经的输入通过发放率对声谱进行编码。使用随机频谱形状(RSS)方法来估计100毫秒时长的频谱平稳刺激的声级在宽频带上如何进行线性和非线性加权。一般来说,发现MNTB神经元及其球状浓密细胞输入可以通过频谱的线性加权得到很好的模拟,这表明通过MNTB的通路能够精确编码声谱,包括那些由头部相关方向传递函数(DTF)提供的声音定位声学线索所产生的声谱。连同MNTB-LSO复合体中用于时间编码的解剖学和生物物理学特性,这些机制可能使人们能够为具有快速频谱时间调制包络的复杂刺激(如语音、动物叫声和移动声源)计算ILD。