Brandenberg Oliver F, Magnus Carsten, Rusert Peter, Regoes Roland R, Trkola Alexandra
Institute of Medical Virology, University of Zürich, Zürich, Switzerland.
Institute of Integrative Biology, ETH Zürich, Zürich, Switzerland.
PLoS Pathog. 2015 Jan 8;11(1):e1004595. doi: 10.1371/journal.ppat.1004595. eCollection 2015 Jan.
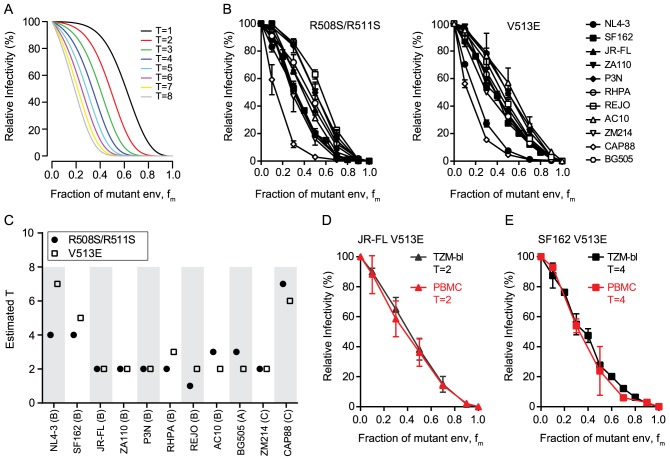
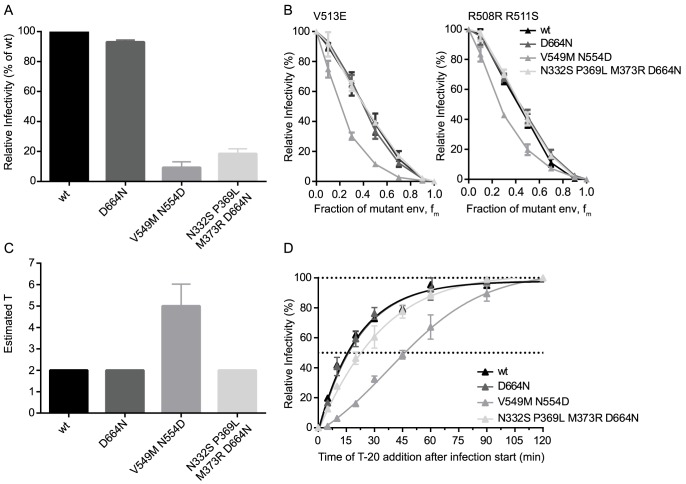
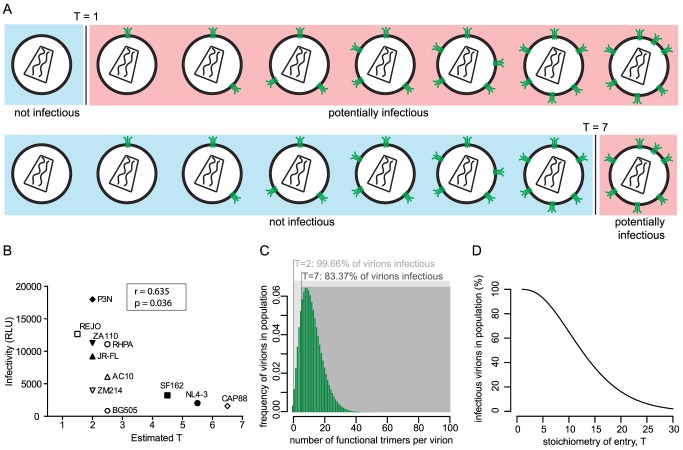
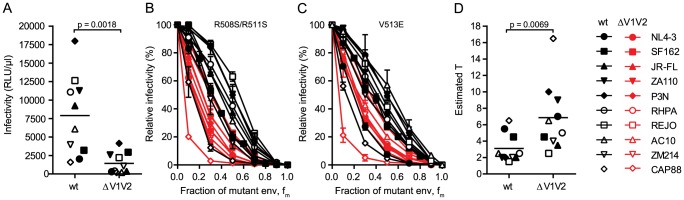
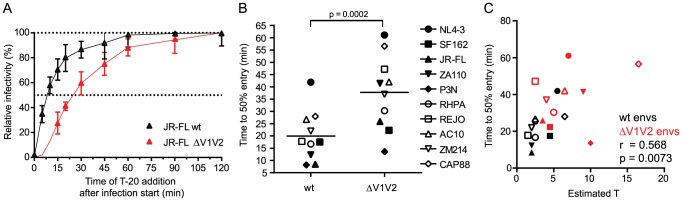
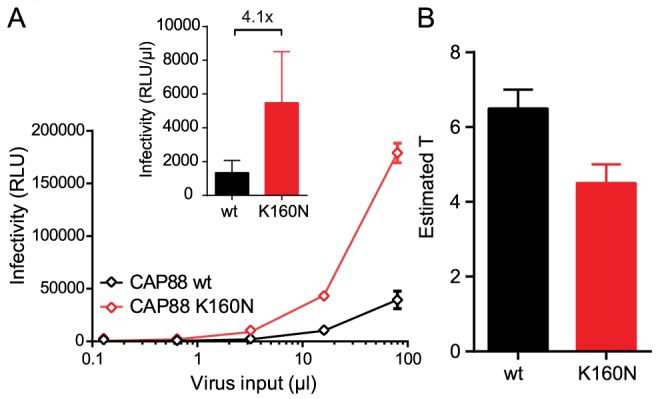
HIV-1 enters target cells by virtue of envelope glycoprotein trimers that are incorporated at low density in the viral membrane. How many trimers are required to interact with target cell receptors to mediate virus entry, the HIV entry stoichiometry, still awaits clarification. Here, we provide estimates of the HIV entry stoichiometry utilizing a combined approach of experimental analyses and mathematical modeling. We demonstrate that divergent HIV strains differ in their stoichiometry of entry and require between 1 to 7 trimers, with most strains depending on 2 to 3 trimers to complete infection. Envelope modifications that perturb trimer structure lead to an increase in the entry stoichiometry, as did naturally occurring antibody or entry inhibitor escape mutations. Highlighting the physiological relevance of our findings, a high entry stoichiometry correlated with low virus infectivity and slow virus entry kinetics. The entry stoichiometry therefore directly influences HIV transmission, as trimer number requirements will dictate the infectivity of virus populations and efficacy of neutralizing antibodies. Thereby our results render consideration of stoichiometric concepts relevant for developing antibody-based vaccines and therapeutics against HIV.
HIV-1借助包膜糖蛋白三聚体进入靶细胞,这些三聚体以低密度整合在病毒膜中。介导病毒进入需要多少三聚体与靶细胞受体相互作用,即HIV进入化学计量比,仍有待阐明。在此,我们利用实验分析和数学建模相结合的方法对HIV进入化学计量比进行了估计。我们证明,不同的HIV毒株进入的化学计量比不同,需要1至7个三聚体,大多数毒株需要2至3个三聚体才能完成感染。扰乱三聚体结构的包膜修饰会导致进入化学计量比增加,天然存在的抗体或进入抑制剂逃逸突变也会如此。我们的发现突出了生理相关性,高进入化学计量比与低病毒感染性和缓慢的病毒进入动力学相关。因此,进入化学计量比直接影响HIV传播,因为三聚体数量需求将决定病毒群体的感染性和中和抗体的效力。由此我们的结果使得在开发针对HIV的基于抗体的疫苗和治疗方法时考虑化学计量学概念变得相关。