Friedlander Tamar, Mayo Avraham E, Tlusty Tsvi, Alon Uri
Department of Molecular Cell Biology, Weizmann Institute of Science, Rehovot, Israel; Institute of Science and Technology Austria, Klosterneuburg, Austria.
Department of Molecular Cell Biology, Weizmann Institute of Science, Rehovot, Israel.
PLoS Comput Biol. 2015 Mar 23;11(3):e1004055. doi: 10.1371/journal.pcbi.1004055. eCollection 2015 Mar.
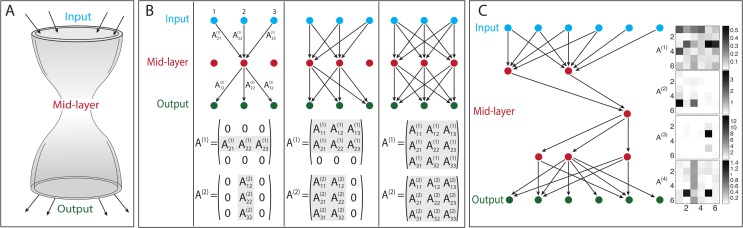
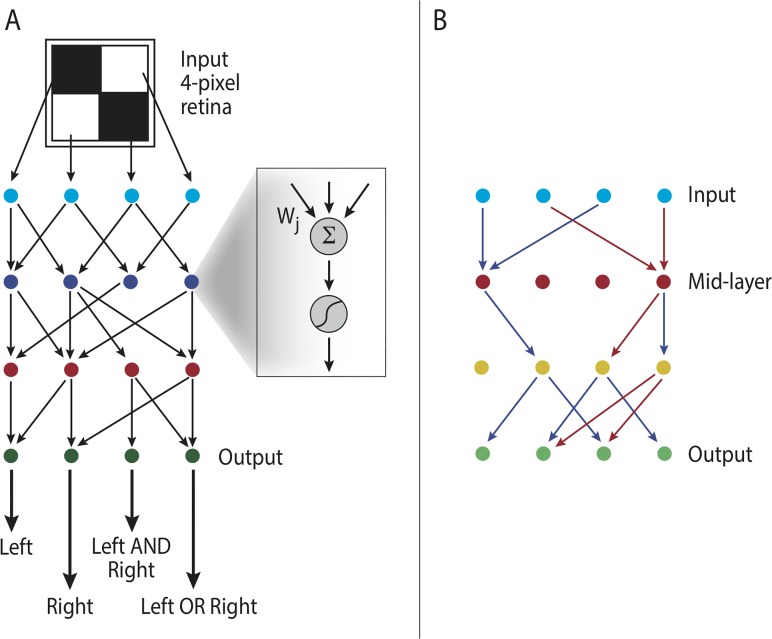
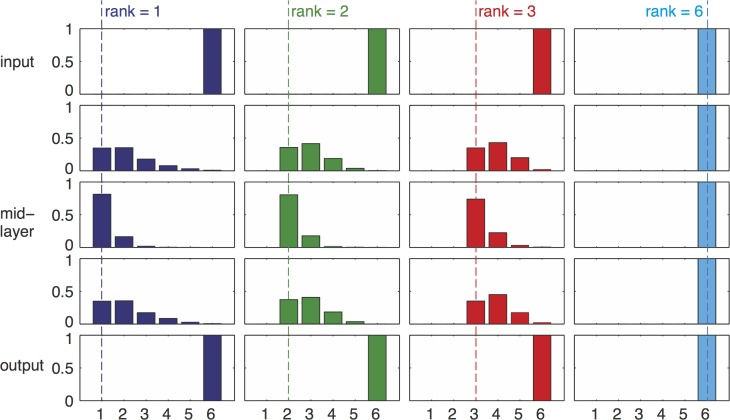
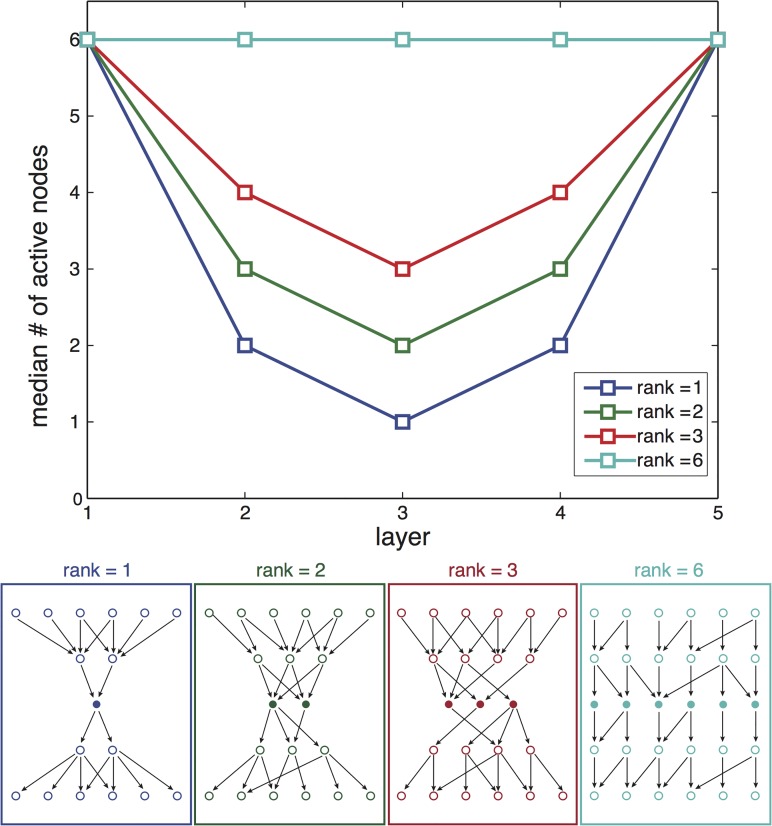
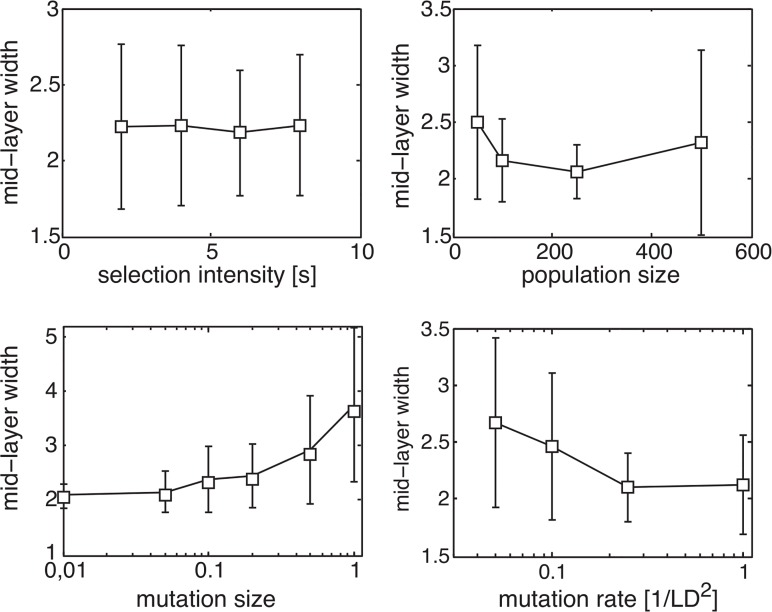
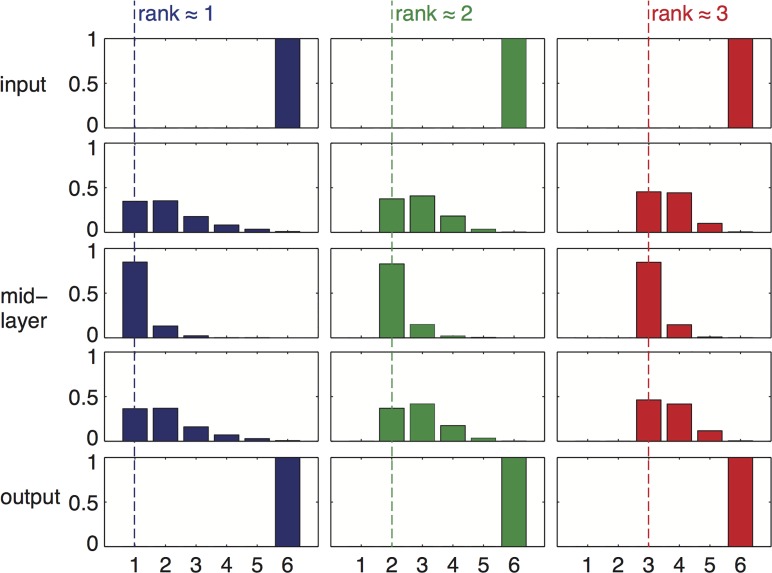
Bow-tie or hourglass structure is a common architectural feature found in many biological systems. A bow-tie in a multi-layered structure occurs when intermediate layers have much fewer components than the input and output layers. Examples include metabolism where a handful of building blocks mediate between multiple input nutrients and multiple output biomass components, and signaling networks where information from numerous receptor types passes through a small set of signaling pathways to regulate multiple output genes. Little is known, however, about how bow-tie architectures evolve. Here, we address the evolution of bow-tie architectures using simulations of multi-layered systems evolving to fulfill a given input-output goal. We find that bow-ties spontaneously evolve when the information in the evolutionary goal can be compressed. Mathematically speaking, bow-ties evolve when the rank of the input-output matrix describing the evolutionary goal is deficient. The maximal compression possible (the rank of the goal) determines the size of the narrowest part of the network-that is the bow-tie. A further requirement is that a process is active to reduce the number of links in the network, such as product-rule mutations, otherwise a non-bow-tie solution is found in the evolutionary simulations. This offers a mechanism to understand a common architectural principle of biological systems, and a way to quantitate the effective rank of the goals under which they evolved.
领结或沙漏结构是许多生物系统中常见的一种结构特征。当多层结构中的中间层所含组件比输入层和输出层少得多时,就会出现领结结构。例如新陈代谢,少数几种构建模块在多种输入营养物质和多种输出生物质成分之间起介导作用;还有信号网络,来自众多受体类型的信息通过一小组信号通路来调节多个输出基因。然而,关于领结结构是如何进化的,人们了解得很少。在这里,我们通过对多层系统进行模拟来实现给定的输入-输出目标,从而探讨领结结构的进化。我们发现,当进化目标中的信息可以被压缩时,领结结构会自发进化。从数学上讲,当描述进化目标的输入-输出矩阵的秩不足时,领结结构就会进化。可能的最大压缩程度(目标的秩)决定了网络最窄部分的大小,也就是领结的大小。另一个要求是有一个过程能够减少网络中的连接数量,比如乘积规则突变,否则在进化模拟中会找到非领结结构的解决方案。这为理解生物系统的一个常见结构原理提供了一种机制,也为量化生物系统进化所依据的目标的有效秩提供了一种方法。