Methot Stephen P, Litzler Ludivine C, Trajtenberg Felipe, Zahn Astrid, Robert Francis, Pelletier Jerry, Buschiazzo Alejandro, Magor Brad G, Di Noia Javier M
Institut de Recherches Cliniques de Montréal, Montréal, Québec H2W 1R7, Canada Department of Medicine, Department of Biochemistry, The Rosalind and Morris Goodman Cancer Research Center, and Department of Oncology, McGill University, Montréal, Québec H3A 0G4, Canada.
Institut de Recherches Cliniques de Montréal, Montréal, Québec H2W 1R7, Canada Department of Biochemistry and Department of Medicine, Université de Montréal, Montréal, Québec H3T 1J4, Canada.
J Exp Med. 2015 Apr 6;212(4):581-96. doi: 10.1084/jem.20141157. Epub 2015 Mar 30.
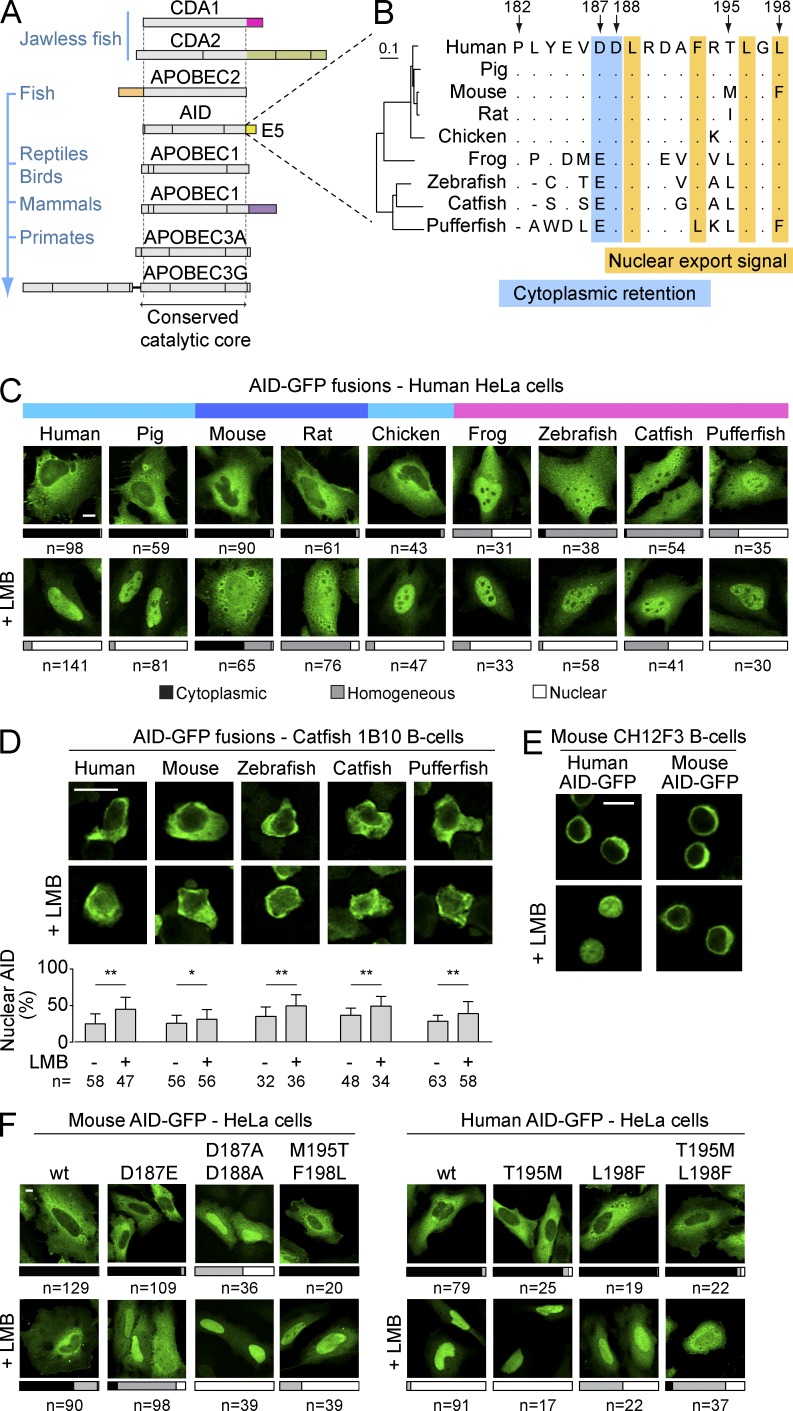
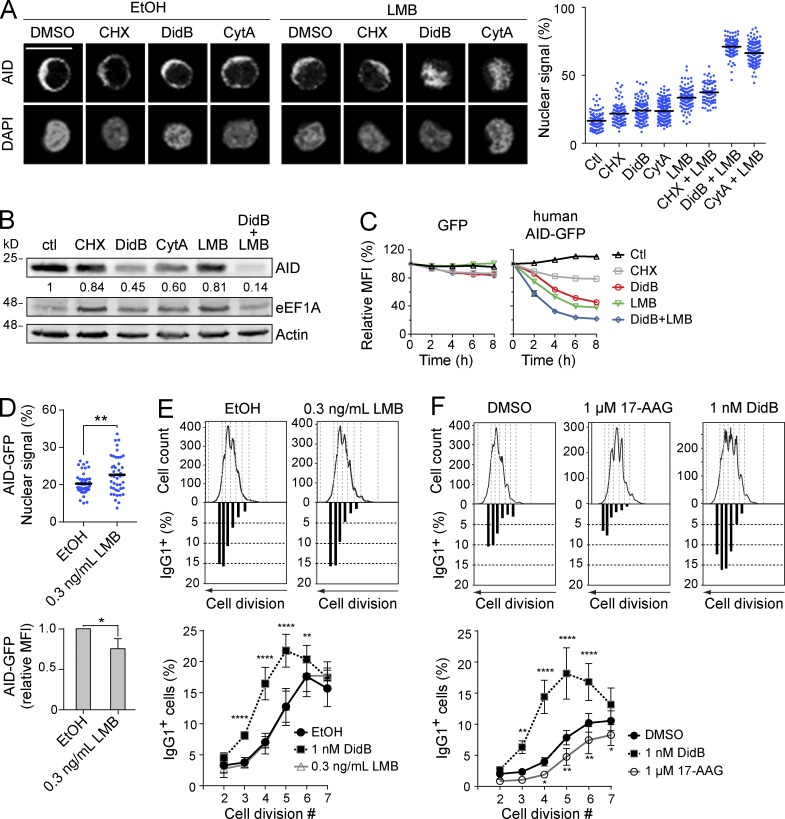
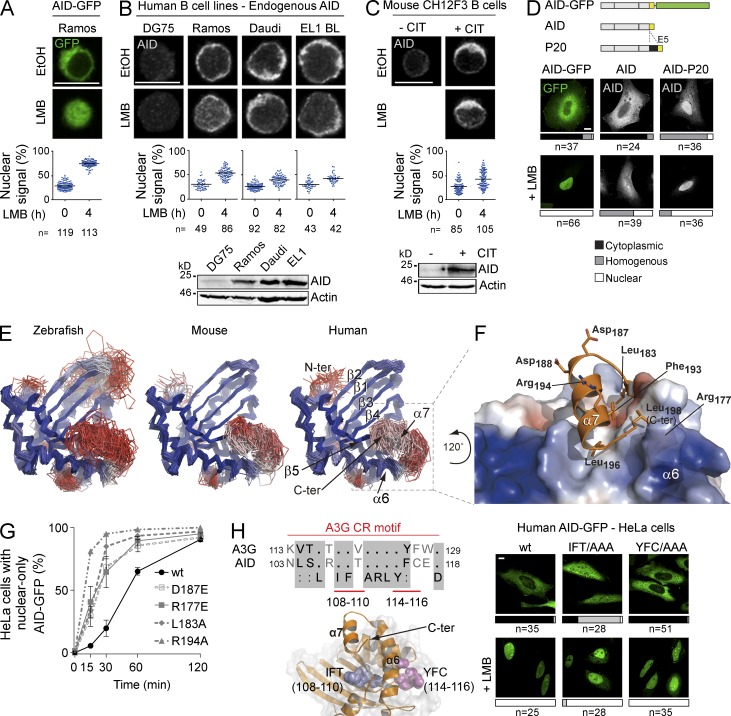
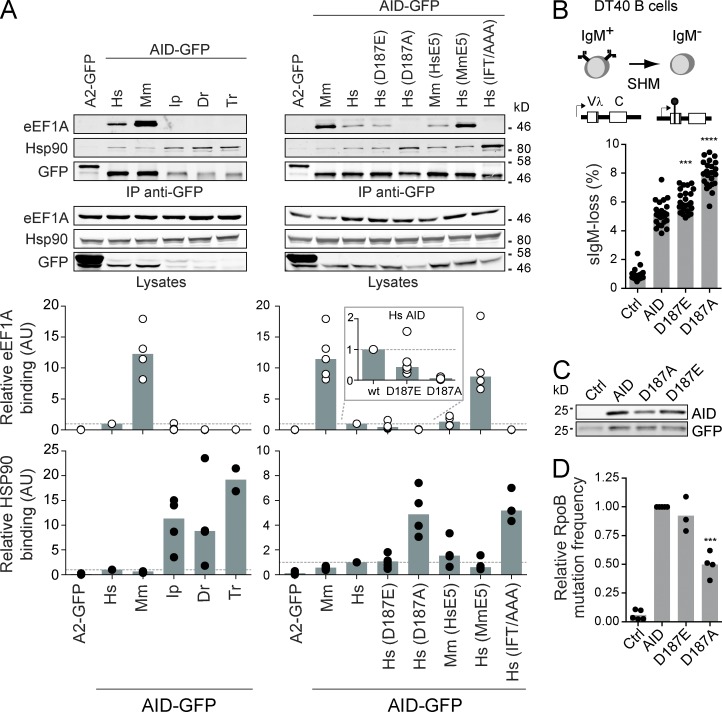
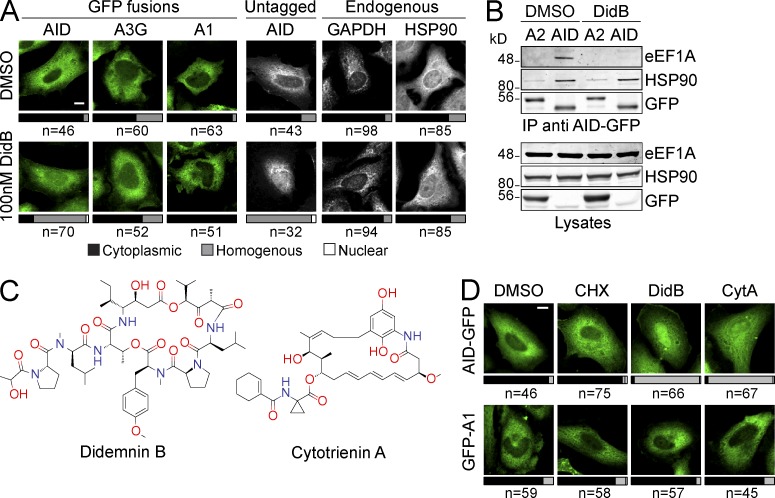
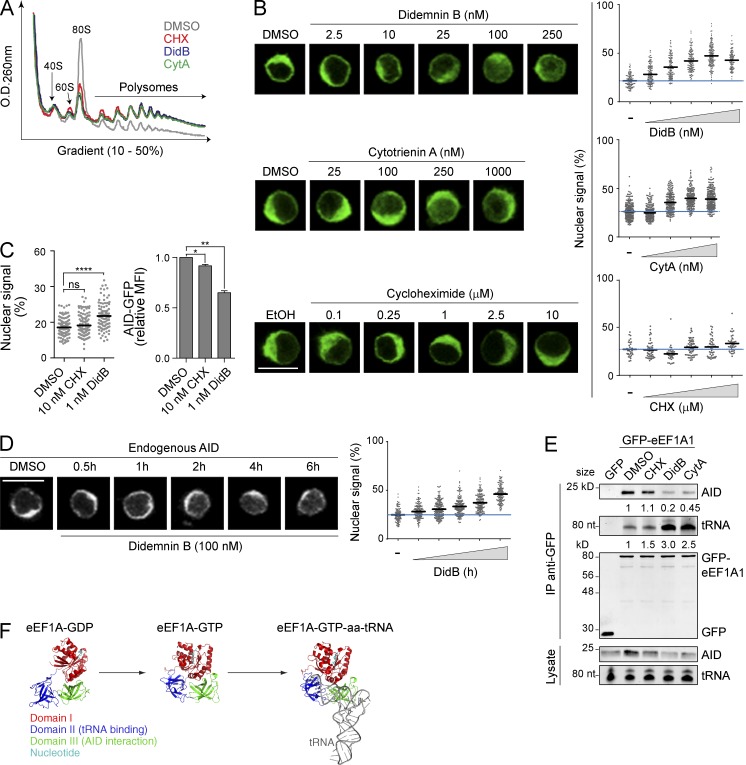
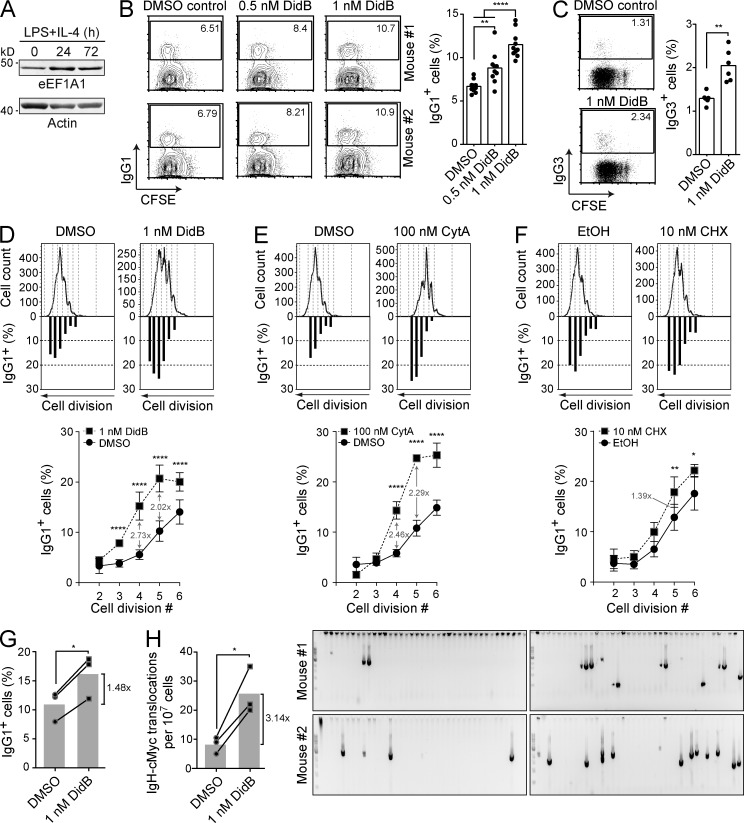
Activation-induced deaminase (AID) initiates mutagenic pathways to diversify the antibody genes during immune responses. The access of AID to the nucleus is limited by CRM1-mediated nuclear export and by an uncharacterized mechanism of cytoplasmic retention. Here, we define a conformational motif in AID that dictates its cytoplasmic retention and demonstrate that the translation elongation factor eukaryotic elongation factor 1 α (eEF1A) is necessary for AID cytoplasmic sequestering. The mechanism is independent of protein synthesis but dependent on a tRNA-free form of eEF1A. Inhibiting eEF1A prevents the interaction with AID, which accumulates in the nucleus and increases class switch recombination as well as chromosomal translocation byproducts. Most AID is associated to unspecified cytoplasmic complexes. We find that the interactions of AID with eEF1A and heat-shock protein 90 kD (HSP90) are inversely correlated. Despite both interactions stabilizing AID, the nature of the AID fractions associated with HSP90 or eEF1A are different, defining two complexes that sequentially produce and store functional AID in the cytoplasm. In addition, nuclear export and cytoplasmic retention cooperate to exclude AID from the nucleus but might not be functionally equivalent. Our results elucidate the molecular basis of AID cytoplasmic retention, define its functional relevance and distinguish it from other mechanisms regulating AID.
激活诱导的胞嘧啶脱氨酶(AID)在免疫反应过程中启动诱变途径,使抗体基因多样化。AID进入细胞核受到CRM1介导的核输出以及一种未知的细胞质滞留机制的限制。在这里,我们确定了AID中一个决定其细胞质滞留的构象基序,并证明翻译延伸因子真核延伸因子1α(eEF1A)是AID细胞质隔离所必需的。该机制独立于蛋白质合成,但依赖于无tRNA形式的eEF1A。抑制eEF1A会阻止其与AID的相互作用,AID会在细胞核中积累,并增加类别转换重组以及染色体易位副产物。大多数AID与未指定的细胞质复合物相关。我们发现AID与eEF1A和热休克蛋白90 kD(HSP90)的相互作用呈负相关。尽管这两种相互作用都能稳定AID,但与HSP90或eEF1A相关的AID组分的性质不同,定义了两种在细胞质中依次产生和储存功能性AID的复合物。此外,核输出和细胞质滞留共同作用将AID排除在细胞核之外,但可能在功能上并不等同。我们的结果阐明了AID细胞质滞留的分子基础,定义了其功能相关性,并将其与其他调节AID的机制区分开来。