Duddy William, Duguez Stephanie, Johnston Helen, Cohen Tatiana V, Phadke Aditi, Gordish-Dressman Heather, Nagaraju Kanneboyina, Gnocchi Viola, Low SiewHui, Partridge Terence
Center for Genetic Medicine Research, Children's National Medical Center, 111 Michigan Avenue NW, Washington DC, 20010 USA ; Myology Center of Research, Institut de Myologie Pitié-Salpétrière - Bâtiment Babinski, 75651 Paris Cedex 13, France.
Center for Genetic Medicine Research, Children's National Medical Center, 111 Michigan Avenue NW, Washington DC, 20010 USA.
Skelet Muscle. 2015 May 1;5:16. doi: 10.1186/s13395-015-0041-y. eCollection 2015.
Preclinical testing of potential therapies for Duchenne muscular dystrophy (DMD) is conducted predominantly of the mdx mouse. But lack of a detailed quantitative description of the pathology of this animal limits our ability to evaluate the effectiveness of putative therapies or their relevance to DMD.
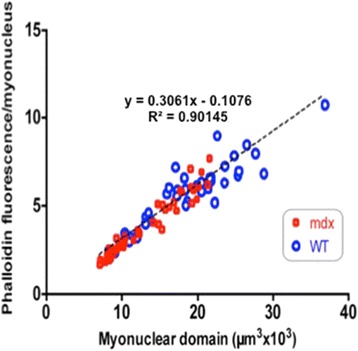
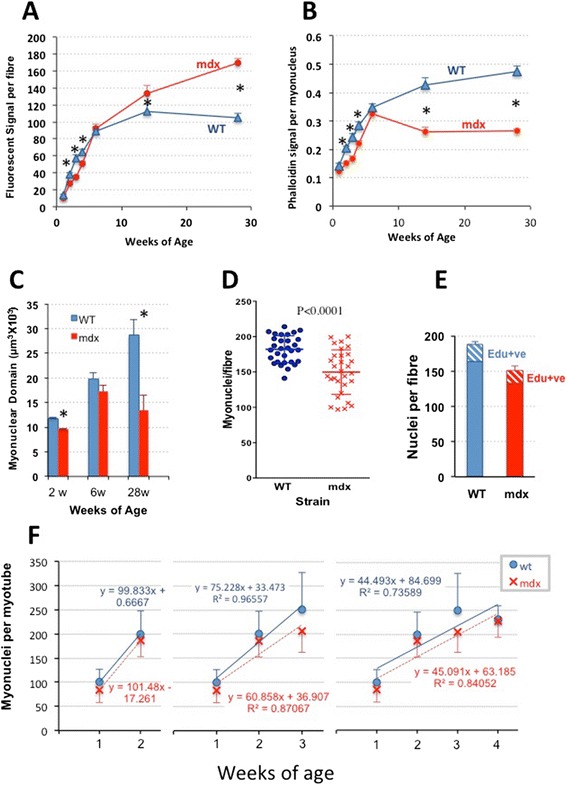
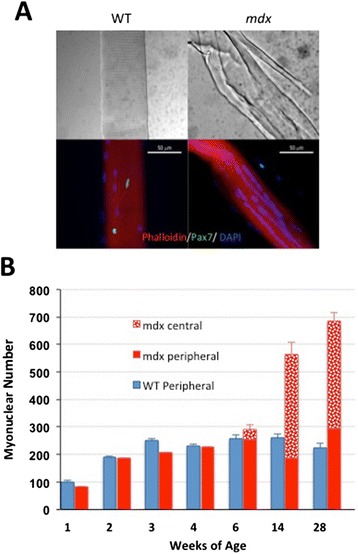
Accordingly, we have measured the main cellular components of muscle growth and regeneration over the period of postnatal growth and early pathology in mdx and wild-type (WT) mice; phalloidin binding is used as a measure of fibre size, myonuclear counts and BrdU labelling as records of myogenic activity.
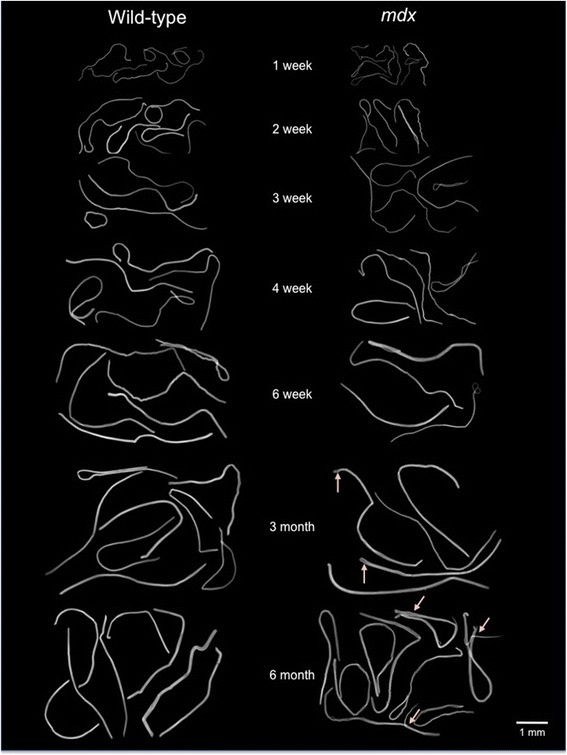
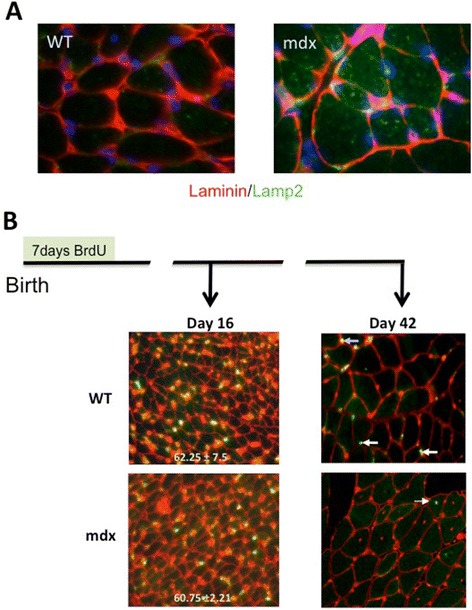
We confirm a two-phase postnatal growth pattern in WT muscle: first, increase in myonuclear number over weeks 1 to 3, then expansion of myonuclear domain. Mdx muscle growth lags behind that of WT prior to overt signs of pathology. Fibres are smaller, with fewer myonuclei and smaller myonuclear domains. Moreover, satellite cells are more readily detached from mdx than WT muscle fibres. At 3 weeks, mdx muscles enter a phase of florid myonecrosis, accompanied by concurrent regeneration of an intensity that results in complete replacement of pre-existing muscle over the succeeding 3 to 4 weeks. Both WT and mdx muscles attain maximum size by 12 to 14 weeks, mdx muscle fibres being up to 50% larger than those of WT as they become increasingly branched. Mdx muscle fibres also become hypernucleated, containing twice as many myonuclei per sarcoplasmic volume, as those of WT, the excess corresponding to the number of centrally placed myonuclei.
The best-known consequence of lack of dystrophin that is common to DMD and the mdx mouse is the conspicuous necrosis and regeneration of muscle fibres. We present protocols for measuring this in terms both of loss of muscle nuclei previously labelled with BrdU and of the intensity of myonuclear labelling with BrdU administered during the regeneration period. Both measurements can be used to assess the efficacy of putative antinecrotic agents. We also show that lack of dystrophin is associated with a number of previously unsuspected abnormalities of muscle fibre structure and function that do not appear to be directly associated with myonecrosis.
杜氏肌营养不良症(DMD)潜在疗法的临床前测试主要在mdx小鼠中进行。但缺乏对该动物病理学的详细定量描述限制了我们评估假定疗法有效性或其与DMD相关性的能力。
因此,我们测量了mdx和野生型(WT)小鼠出生后生长和早期病理学期间肌肉生长和再生的主要细胞成分;鬼笔环肽结合用于测量纤维大小,肌核计数和BrdU标记作为肌源性活动的记录。
我们证实了WT肌肉出生后有两阶段生长模式:首先,在第1至3周肌核数量增加,然后是肌核域的扩展。在出现明显病理学迹象之前,mdx肌肉生长落后于WT。纤维较小,肌核较少且肌核域较小。此外,卫星细胞比WT肌纤维更容易从mdx肌纤维上脱离。在3周时,mdx肌肉进入明显的肌坏死阶段,同时伴有强度相当的再生,导致在接下来的3至4周内完全替换先前存在的肌肉。WT和mdx肌肉在12至14周时达到最大尺寸,mdx肌纤维在变得越来越分支时比WT肌纤维大50%。mdx肌纤维也变得多核化,每肌浆体积中的肌核数量是WT的两倍,多余的部分对应于中央放置的肌核数量。
DMD和mdx小鼠共有的缺乏抗肌萎缩蛋白最著名的后果是肌纤维明显的坏死和再生。我们提出了根据先前用BrdU标记的肌核损失以及再生期给予BrdU后肌核标记强度来测量此情况的方案。这两种测量都可用于评估假定的抗坏死剂的疗效。我们还表明,缺乏抗肌萎缩蛋白与许多先前未被怀疑的肌纤维结构和功能异常有关,这些异常似乎与肌坏死没有直接关联。