Gamisch Alexander, Fischer Gunter Alexander, Comes Hans Peter
Department of Ecology and Evolution, University of Salzburg, A-5020, Salzburg, Austria.
Kadoorie Farm and Botanic Garden Corporation, Lam Kam Road, Tai Po, NT, Hong Kong.
BMC Evol Biol. 2015 Sep 16;15:192. doi: 10.1186/s12862-015-0471-5.
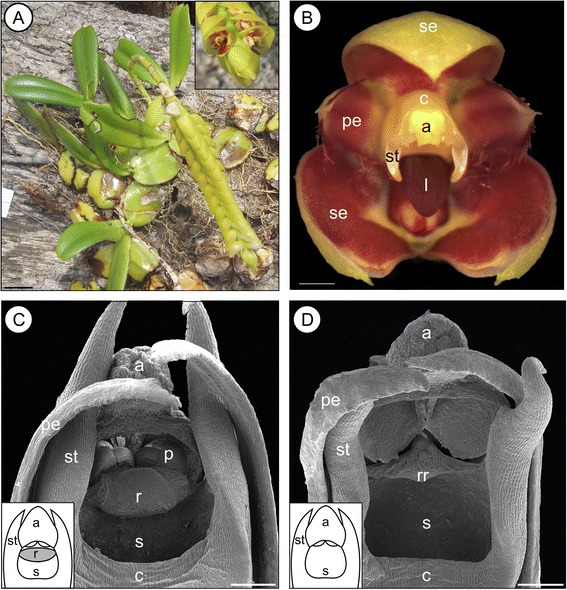
The transition from outcrossing to selfing has long been portrayed as an 'evolutionary dead end' because, first, reversals are unlikely and, second, selfing lineages suffer from higher rates of extinction owing to a reduced potential for adaptation and the accumulation of deleterious mutations. We tested these two predictions in a clade of Madagascan Bulbophyllum orchids (30 spp.), including eight species where auto-pollinating morphs (i.e., selfers, without a 'rostellum') co-exist with their pollinator-dependent conspecifics (i.e., outcrossers, possessing a rostellum). Specifically, we addressed this issue on the basis of a time-calibrated phylogeny by means of ancestral character reconstructions and within the state-dependent evolution framework of BiSSE (Binary State Speciation and Extinction), which allowed jointly estimating rates of transition, speciation, and extinction between outcrossing and selfing.
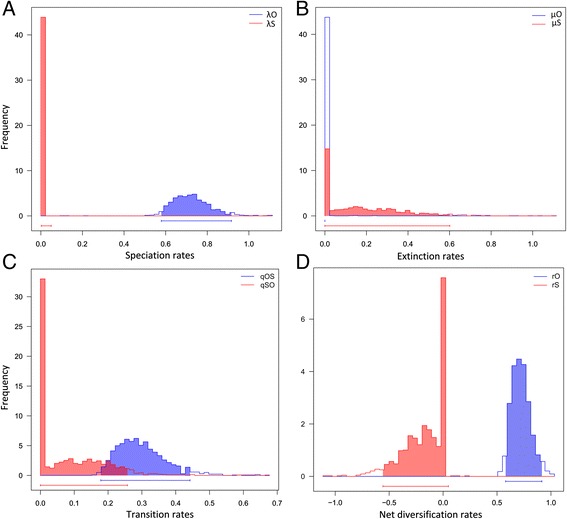
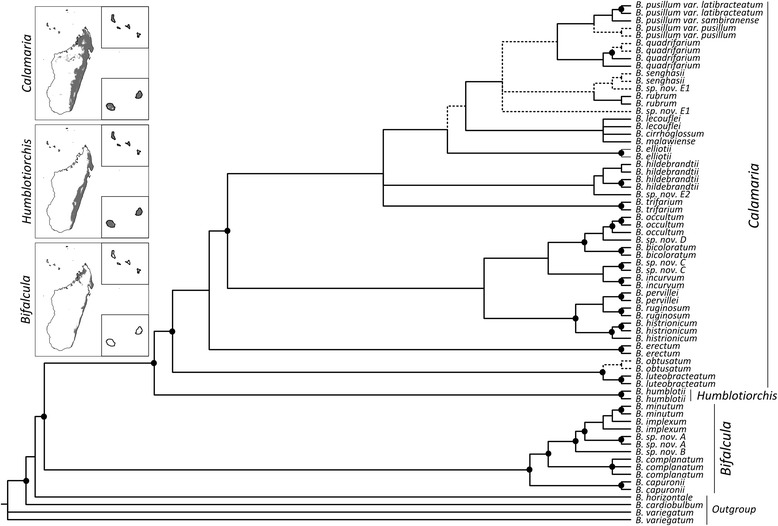
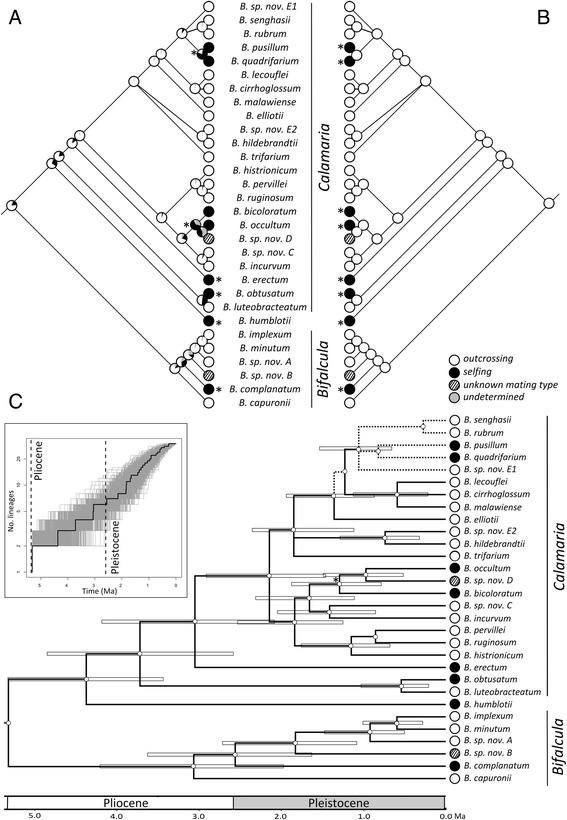
The eight species capable of selfing occurred in scattered positions across the phylogeny, with two likely originating in the Pliocene (ca. 4.4-3.1 Ma), one in the Early Pleistocene (ca. 2.4 Ma), and five since the mid-Pleistocene (ca. ≤ 1.3 Ma). We infer that this scattered phylogenetic distribution of selfing is best described by models including up to eight independent outcrossing-to-selfing transitions and very low rates of speciation (and either moderate or zero rates of extinction) associated with selfing.
The frequent and irreversible outcrossing-to-selfing transitions in Madagascan Bulbophyllum are clearly congruent with the first prediction of the dead end hypothesis. The inability of our study to conclusively reject or support the likewise predicted higher extinction rate in selfing lineages might be explained by a combination of methodological limitations (low statistical power of our BiSSE approach to reliably estimate extinction in small-sized trees) and evolutionary processes (insufficient time elapsed for selfers to go extinct). We suggest that, in these tropical orchids, a simple genetic basis of selfing (via loss of the 'rostellum') is needed to explain the strikingly recurrent transitions to selfing, perhaps reflecting rapid response to parallel and novel selective environments over Late Quaternary (≤ 1.3 Ma) time scales.
从异交向自交的转变长期以来一直被描绘为一个“进化死胡同”,原因如下:其一,逆转不太可能发生;其二,自交谱系由于适应潜力降低和有害突变的积累,灭绝率更高。我们在马达加斯加石豆兰属兰花的一个分支(30个物种)中检验了这两个预测,其中包括8个物种,在这些物种中,自动授粉形态(即自交者,没有“蕊喙”)与其依赖传粉者的同种形态(即异交者,拥有蕊喙)共存。具体而言,我们在一个经过时间校准的系统发育树上,通过祖先性状重建,并在BiSSE(二元状态物种形成和灭绝)的状态依赖进化框架内解决了这个问题,该框架允许联合估计异交和自交之间的转变、物种形成和灭绝速率。
能够自交的8个物种在系统发育树上分布零散,其中两个可能起源于上新世(约440 - 310万年前),一个起源于早更新世(约240万年前),还有5个自更新世中期以来(约≤130万年前)。我们推断,这种自交的零散系统发育分布最好用包含多达8次独立的异交向自交转变以及与自交相关的极低物种形成速率(以及中等或零灭绝速率)的模型来描述。
马达加斯加石豆兰中频繁且不可逆的异交向自交转变显然与“进化死胡同”假说的第一个预测相符。我们的研究无法确凿地拒绝或支持同样预测的自交谱系中更高灭绝率,这可能是由于方法学限制(我们的BiSSE方法在估计小型系统发育树上的灭绝时统计能力较低)和进化过程(自交者灭绝所需的时间不足)共同导致的。我们认为,在这些热带兰花中,自交(通过“蕊喙”的丧失)的简单遗传基础是解释向自交显著反复转变的必要条件,这或许反映了在晚更新世(≤130万年前)时间尺度上对平行和新选择环境的快速响应。