Thorley-Lawson David A
School of Medicine, Tufts University, Boston, MA, 02111, USA.
Curr Top Microbiol Immunol. 2015;390(Pt 1):151-209. doi: 10.1007/978-3-319-22822-8_8.
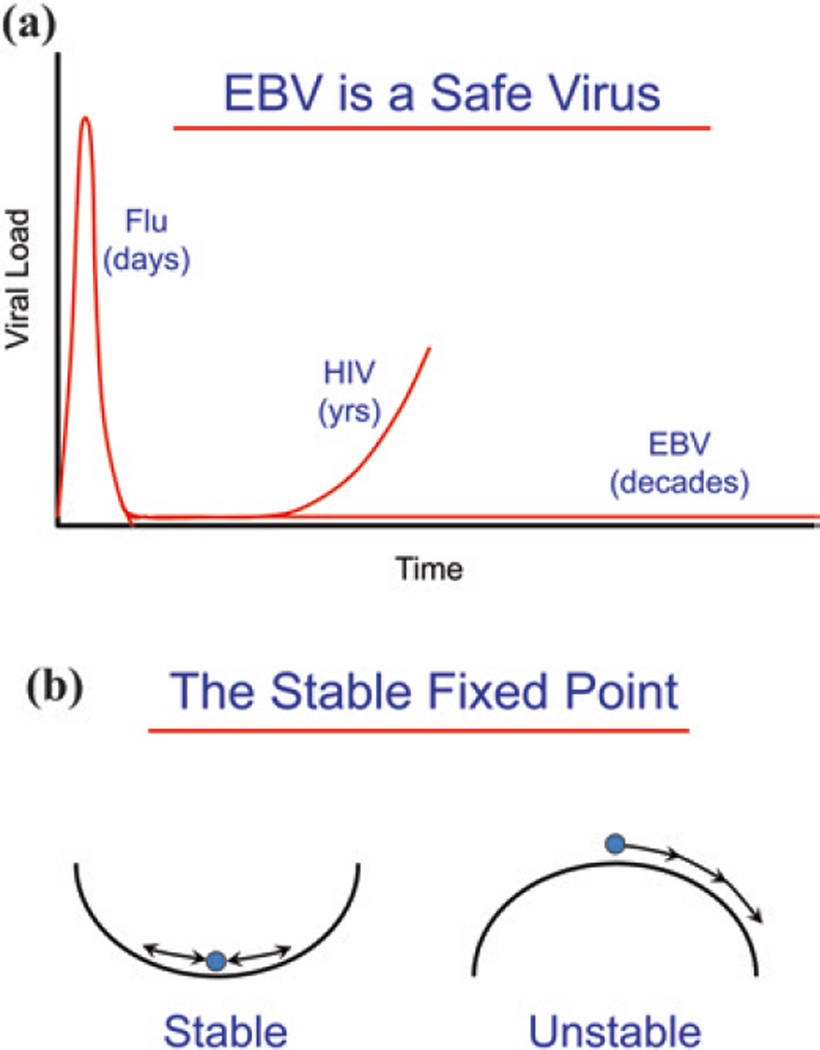
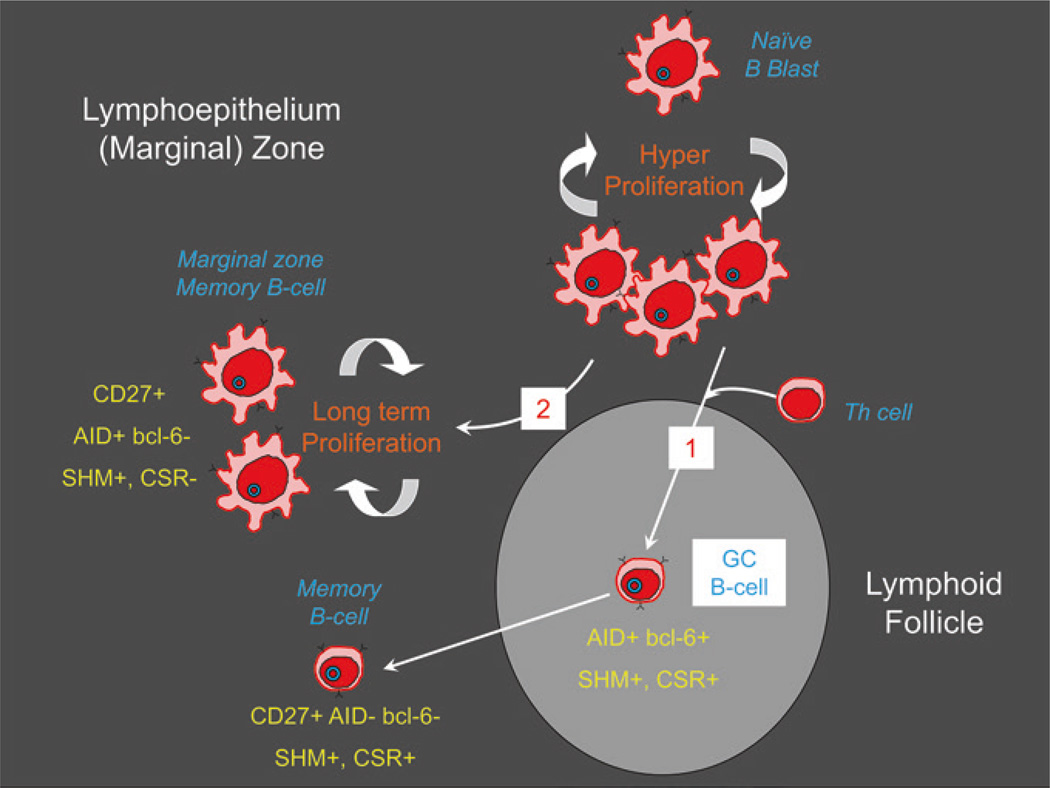
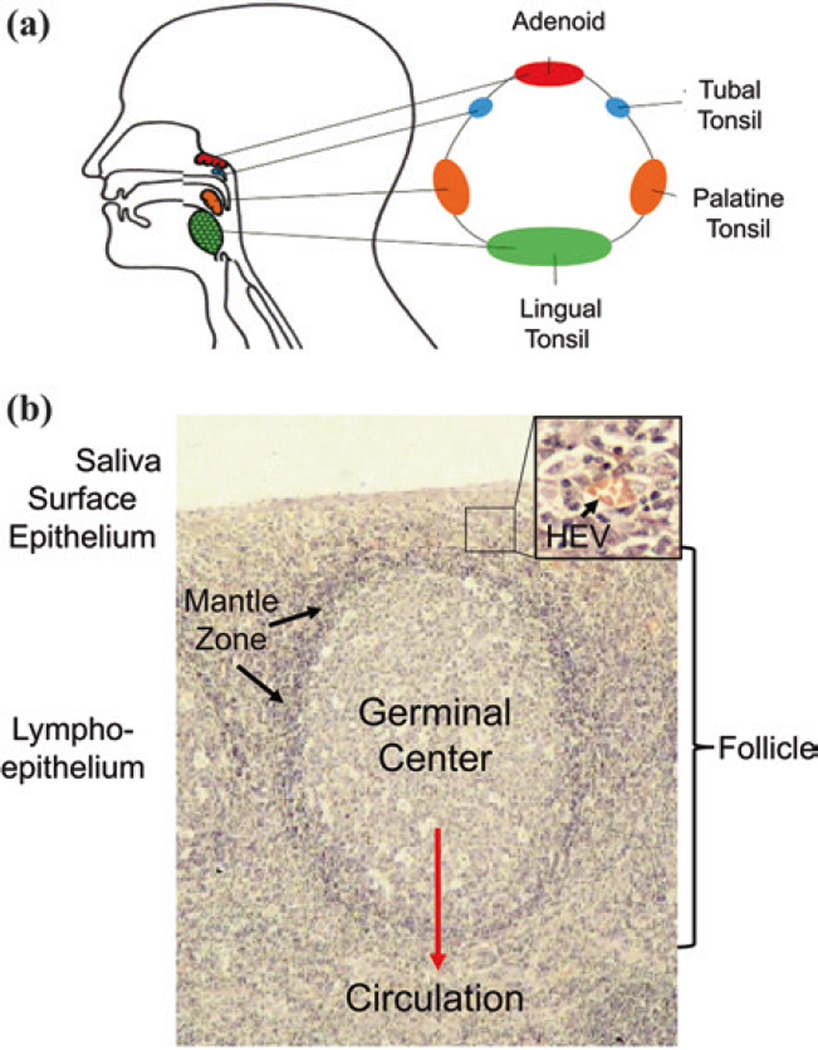
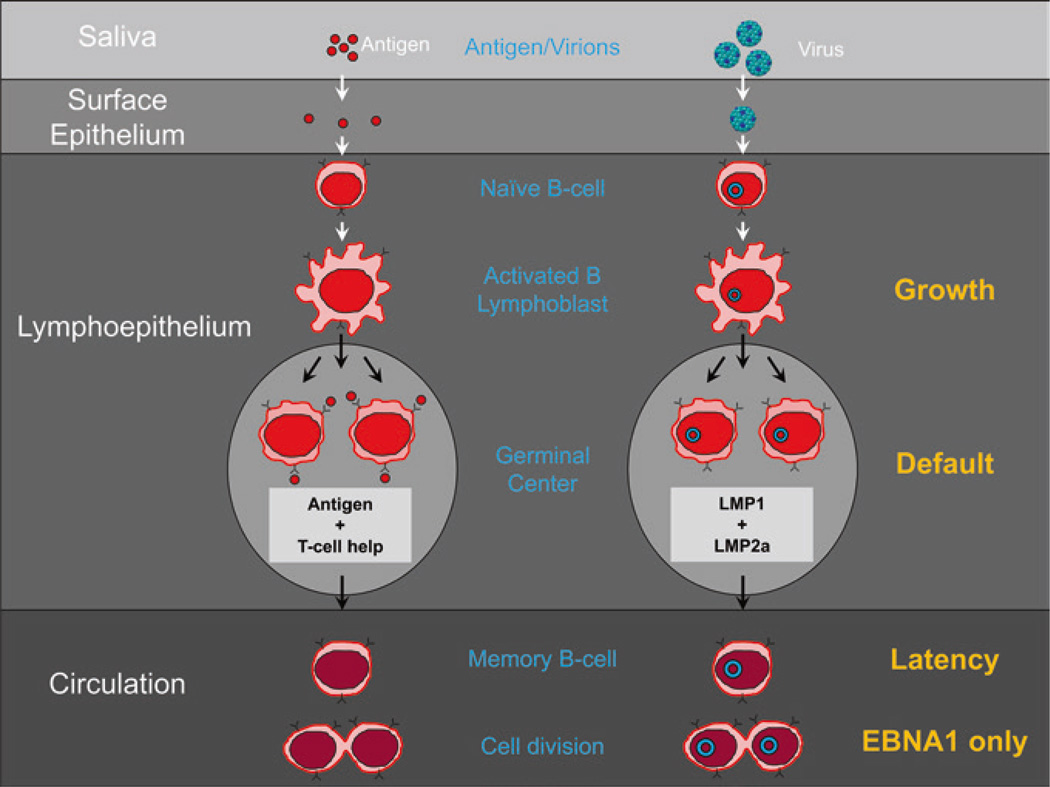
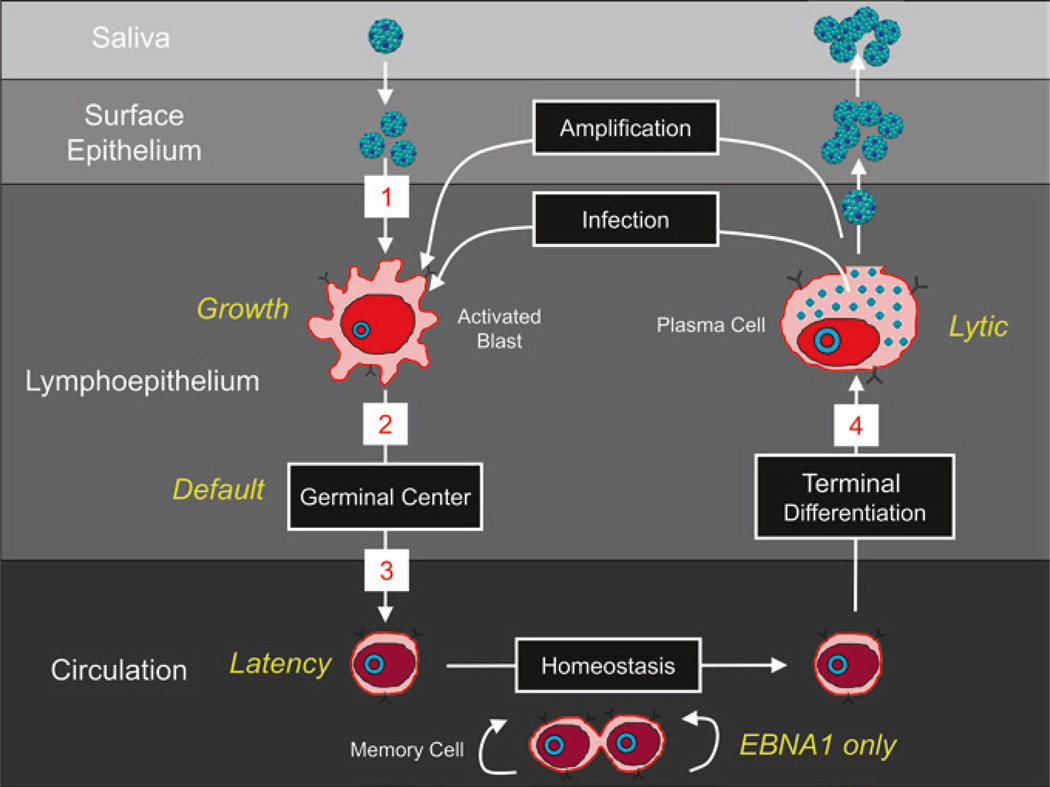
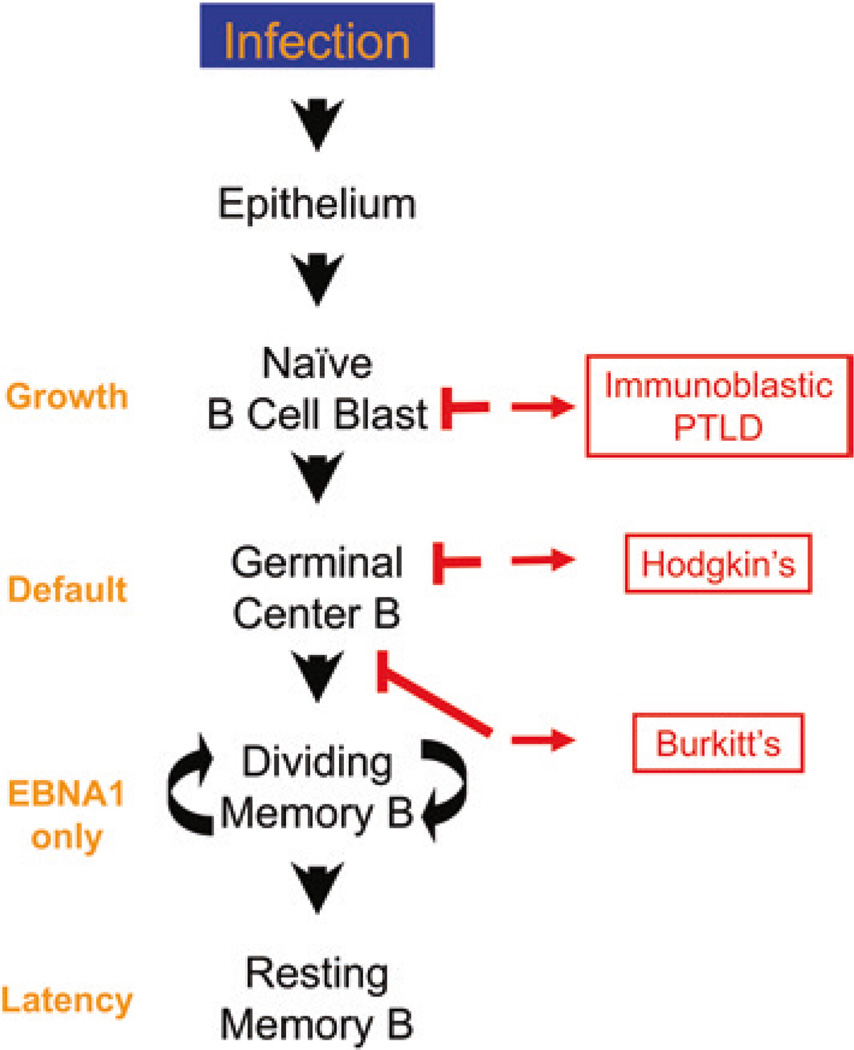
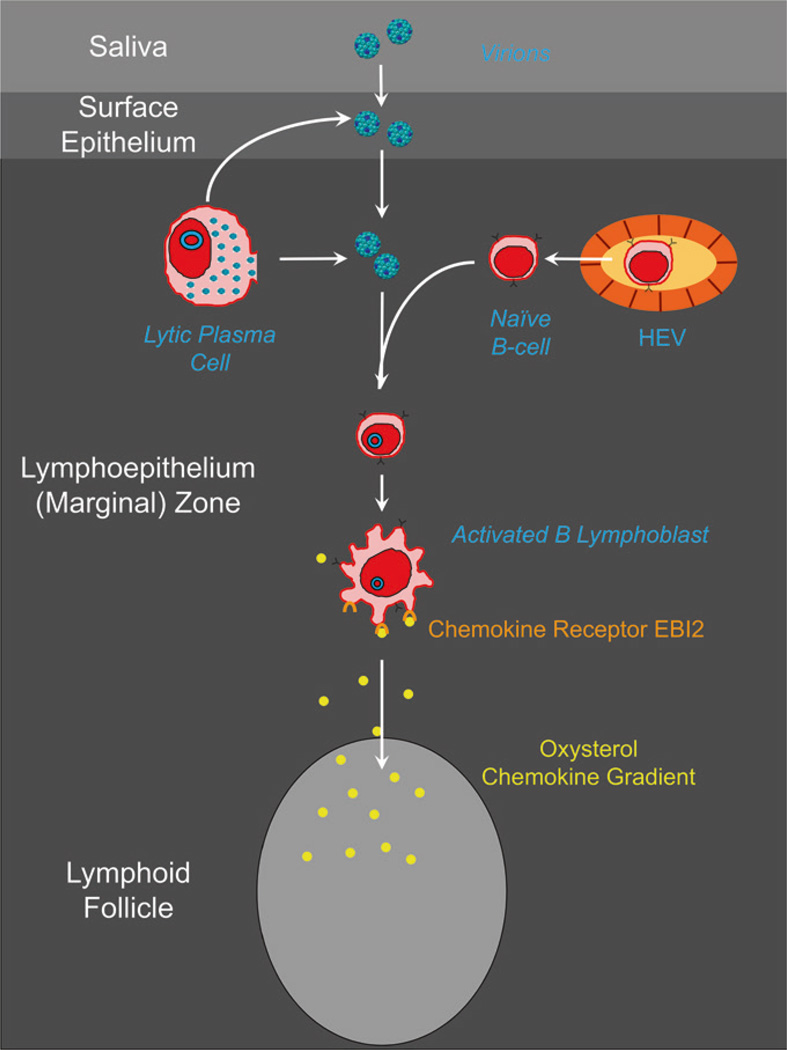
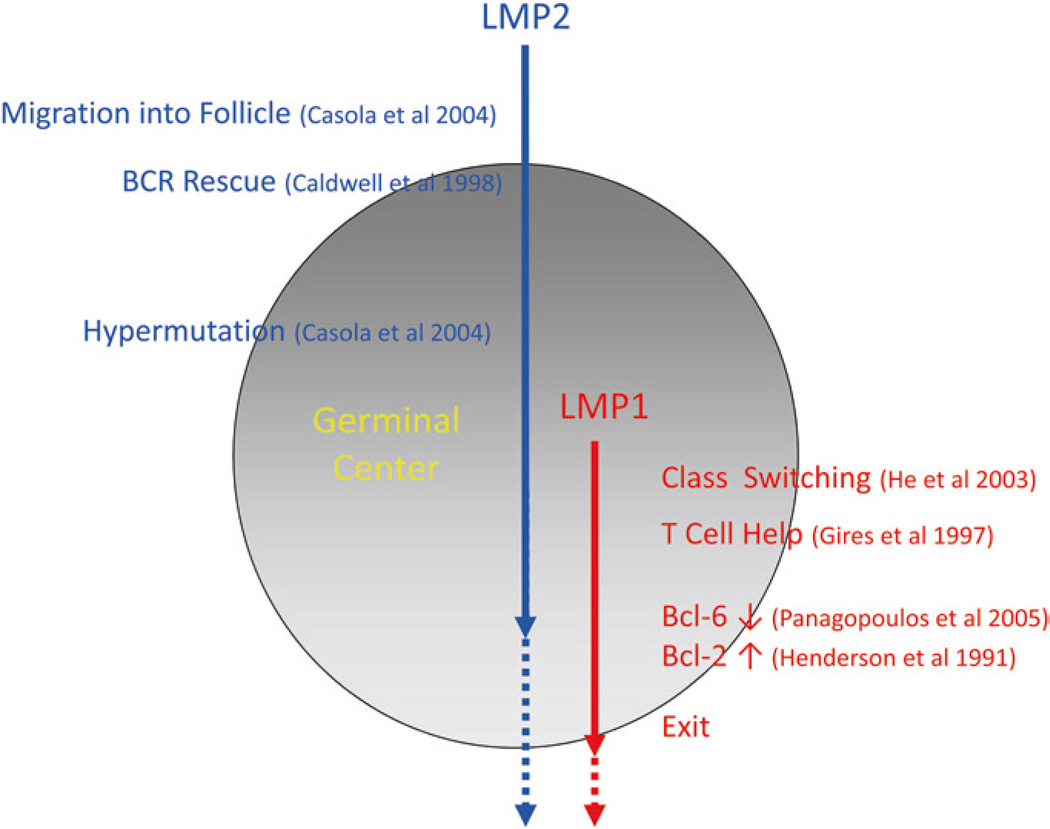
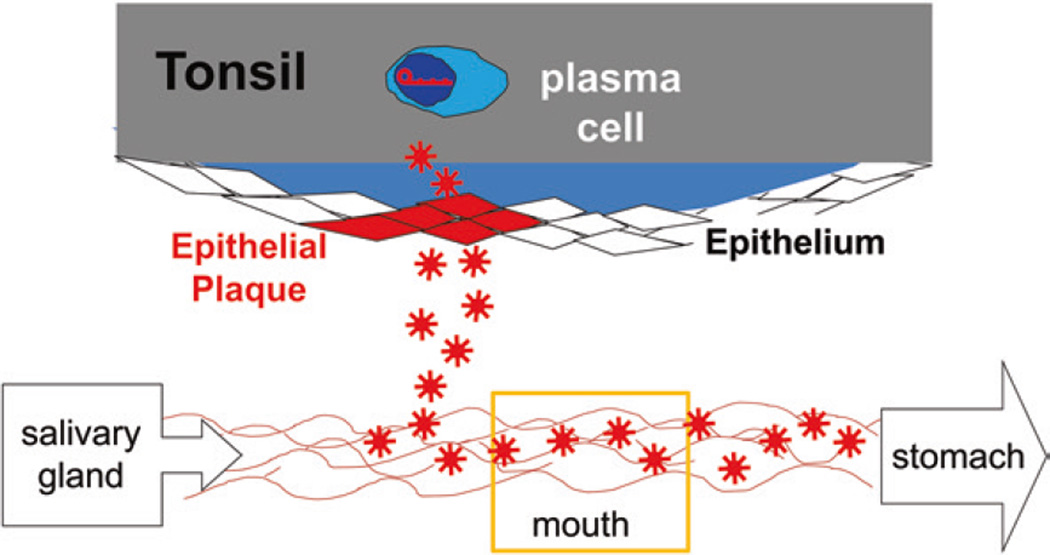
Persistent infection by EBV is explained by the germinal center model (GCM) which provides a satisfying and currently the only explanation for EBVs disparate biology. Since the GCM touches on every aspect of the virus, this chapter will serve as an introduction to the subsequent chapters. EBV is B lymphotropic, and its biology closely follows that of normal mature B lymphocytes. The virus persists quiescently in resting memory B cells for the lifetime of the host in a non-pathogenic state that is also invisible to the immune response. To access this compartment, the virus infects naïve B cells in the lymphoepithelium of the tonsils and activates these cells using the growth transcription program. These cells migrate to the GC where they switch to a more limited transcription program, the default program, which helps rescue them into the memory compartment where the virus persists. For egress, the infected memory cells return to the lymphoepithelium where they occasionally differentiate into plasma cells activating viral replication. The released virus can either infect more naïve B cells or be amplified in the epithelium for shedding. This cycle of infection and the quiescent state in memory B cells allow for lifetime persistence at a very low level that is remarkably stable over time. Mathematically, this is a stable fixed point where the mechanisms regulating persistence drive the state back to equilibrium when perturbed. This is the GCM of EBV persistence. Other possible sites and mechanisms of persistence will also be discussed.
EB病毒的持续感染可以用生发中心模型(GCM)来解释,该模型为EB病毒截然不同的生物学特性提供了令人满意且目前唯一的解释。由于生发中心模型涉及病毒的各个方面,本章将作为后续章节的引言。EB病毒具有嗜B淋巴细胞性,其生物学特性与正常成熟B淋巴细胞密切相关。在宿主的一生中,该病毒以一种非致病性状态在静止的记忆B细胞中静止持续存在,这种状态也不会引起免疫反应。为了进入这个细胞区室,病毒感染扁桃体淋巴上皮中的初始B细胞,并利用生长转录程序激活这些细胞。这些细胞迁移到生发中心,在那里它们切换到一个更有限的转录程序,即默认程序,这有助于它们进入病毒持续存在的记忆细胞区室。为了释放,被感染的记忆细胞回到淋巴上皮,在那里它们偶尔分化为浆细胞,从而激活病毒复制。释放出的病毒要么感染更多的初始B细胞,要么在上皮细胞中扩增以便排出。这种感染循环以及记忆B细胞中的静止状态使得病毒能够在非常低的水平上持续终身,而且随着时间的推移非常稳定。从数学角度来看,这是一个稳定的固定点,调节持续存在的机制在受到干扰时会将状态推回到平衡。这就是EB病毒持续存在的生发中心模型。还将讨论其他可能的持续存在位点和机制。