Absmeier Eva, Wollenhaupt Jan, Mozaffari-Jovin Sina, Becke Christian, Lee Chung-Tien, Preussner Marco, Heyd Florian, Urlaub Henning, Lührmann Reinhard, Santos Karine F, Wahl Markus C
Laboratory of Structural Biochemistry, Freie Universität Berlin, D-14195 Berlin, Germany;
Department of Cellular Biochemistry, Max Planck Institute for Biophysical Chemistry, D-37077 Göttingen, Germany;
Genes Dev. 2015 Dec 15;29(24):2576-87. doi: 10.1101/gad.271528.115. Epub 2015 Dec 4.
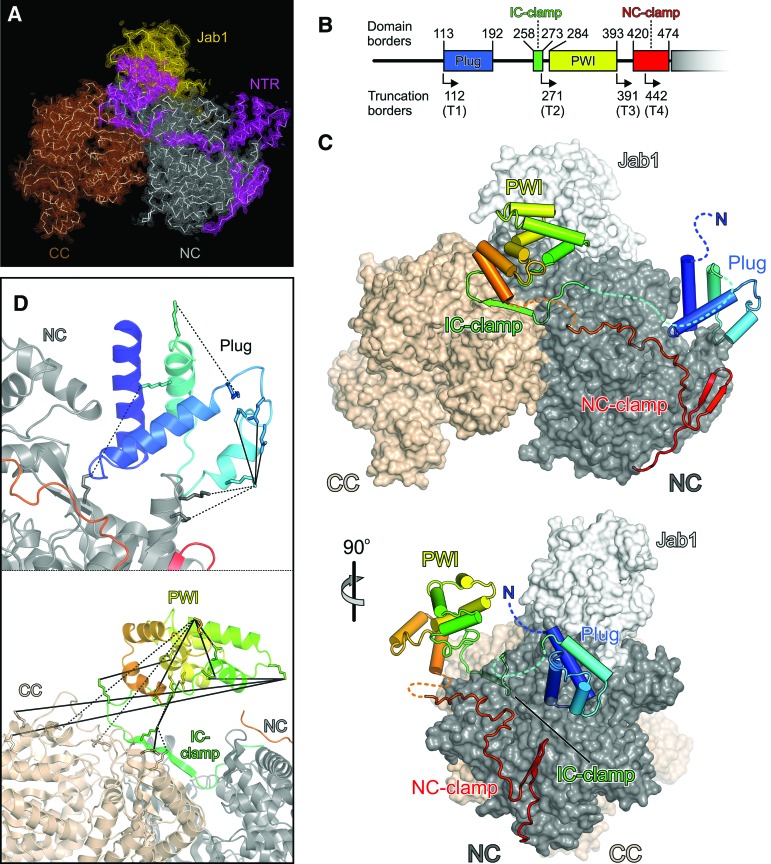
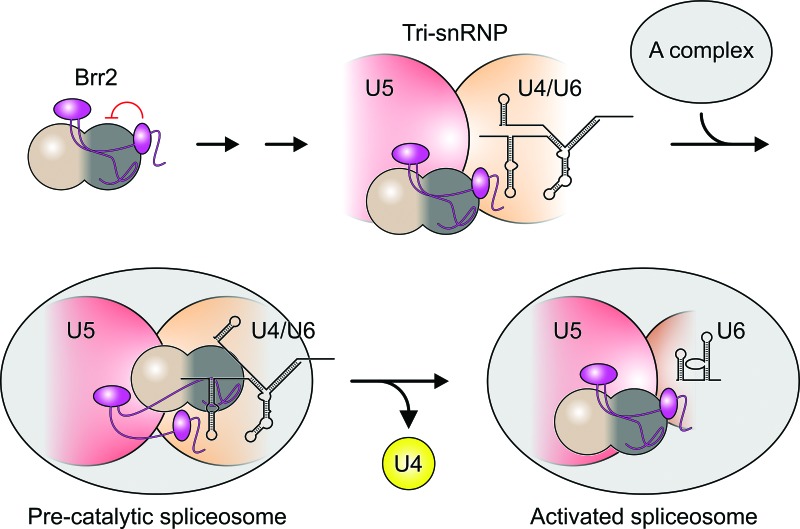
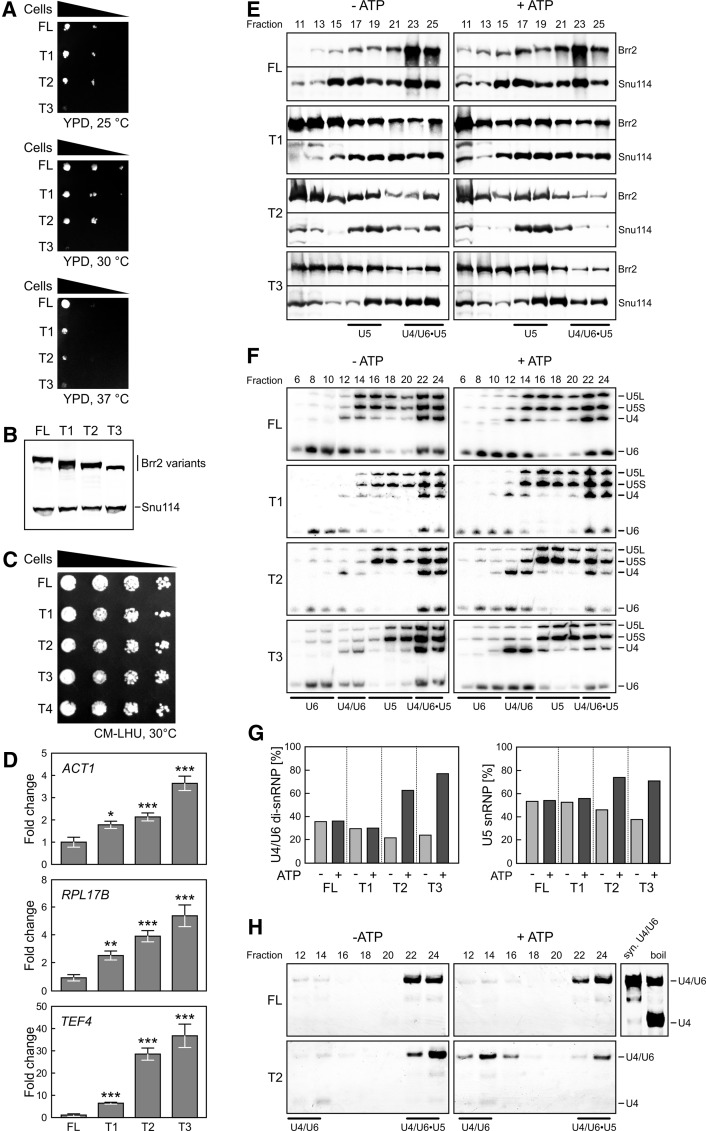
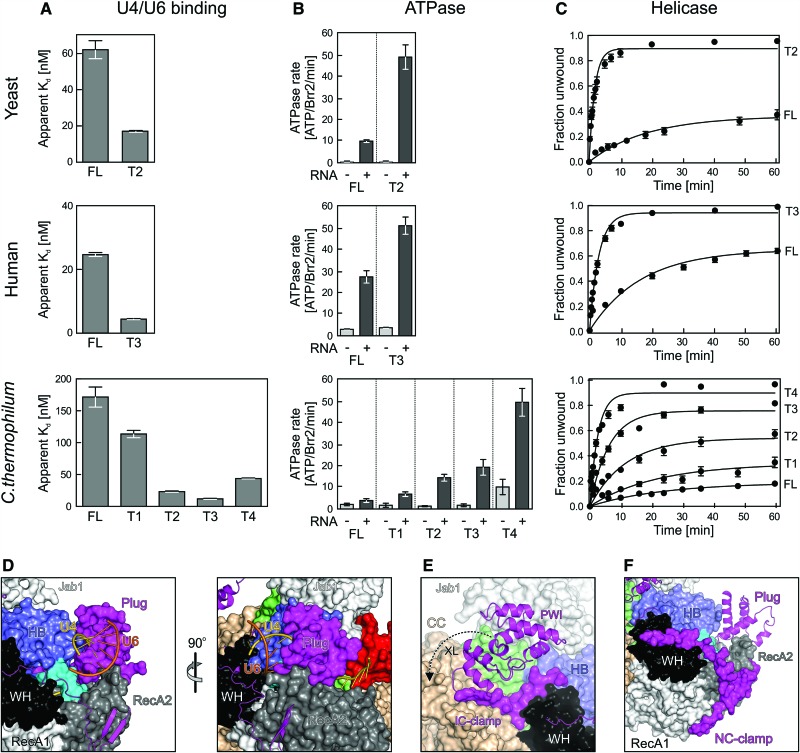
The Brr2 helicase provides the key remodeling activity for spliceosome catalytic activation, during which it disrupts the U4/U6 di-snRNP (small nuclear RNA protein), and its activity has to be tightly regulated. Brr2 exhibits an unusual architecture, including an ∼ 500-residue N-terminal region, whose functions and molecular mechanisms are presently unknown, followed by a tandem array of structurally similar helicase units (cassettes), only the first of which is catalytically active. Here, we show by crystal structure analysis of full-length Brr2 in complex with a regulatory Jab1/MPN domain of the Prp8 protein and by cross-linking/mass spectrometry of isolated Brr2 that the Brr2 N-terminal region encompasses two folded domains and adjacent linear elements that clamp and interconnect the helicase cassettes. Stepwise N-terminal truncations led to yeast growth and splicing defects, reduced Brr2 association with U4/U6•U5 tri-snRNPs, and increased ATP-dependent disruption of the tri-snRNP, yielding U4/U6 di-snRNP and U5 snRNP. Trends in the RNA-binding, ATPase, and helicase activities of the Brr2 truncation variants are fully rationalized by the crystal structure, demonstrating that the N-terminal region autoinhibits Brr2 via substrate competition and conformational clamping. Our results reveal molecular mechanisms that prevent premature and unproductive tri-snRNP disruption and suggest novel principles of Brr2-dependent splicing regulation.
Brr2解旋酶为剪接体的催化激活提供关键的重塑活性,在此过程中它会破坏U4/U6二小分子核糖核蛋白(small nuclear RNA protein, snRNP),其活性必须受到严格调控。Brr2具有不同寻常的结构,包括一个约500个残基的N端区域,其功能和分子机制目前尚不清楚,随后是一系列结构相似的解旋酶单元(模块)串联排列,其中只有第一个具有催化活性。在这里,我们通过对与Prp8蛋白的调控Jab1/MPN结构域结合的全长Brr2进行晶体结构分析,以及对分离的Brr2进行交联/质谱分析表明,Brr2的N端区域包含两个折叠结构域和相邻的线性元件,它们夹住并连接解旋酶模块。逐步进行N端截短导致酵母生长和剪接缺陷,减少Brr2与U4/U6•U5三小分子核糖核蛋白(tri-snRNPs)的结合,并增加ATP依赖的三小分子核糖核蛋白的破坏,产生U4/U6二小分子核糖核蛋白和U5小分子核糖核蛋白。Brr2截短变体的RNA结合、ATP酶和解旋酶活性趋势通过晶体结构得到了充分的解释,表明N端区域通过底物竞争和构象钳制对Brr2进行自抑制。我们的结果揭示了防止三小分子核糖核蛋白过早和无效破坏的分子机制,并提出了Brr2依赖性剪接调控的新原理。