Diz-Muñoz Alba, Thurley Kevin, Chintamen Sana, Altschuler Steven J, Wu Lani F, Fletcher Daniel A, Weiner Orion D
Cardiovascular Research Institute and Department of Biochemistry and Biophysics, University of California San Francisco, San Francisco, California, United States of America.
Bioengineering Department and Biophysics Program, University of California Berkeley, Berkeley, California, United States of America.
PLoS Biol. 2016 Jun 9;14(6):e1002474. doi: 10.1371/journal.pbio.1002474. eCollection 2016 Jun.
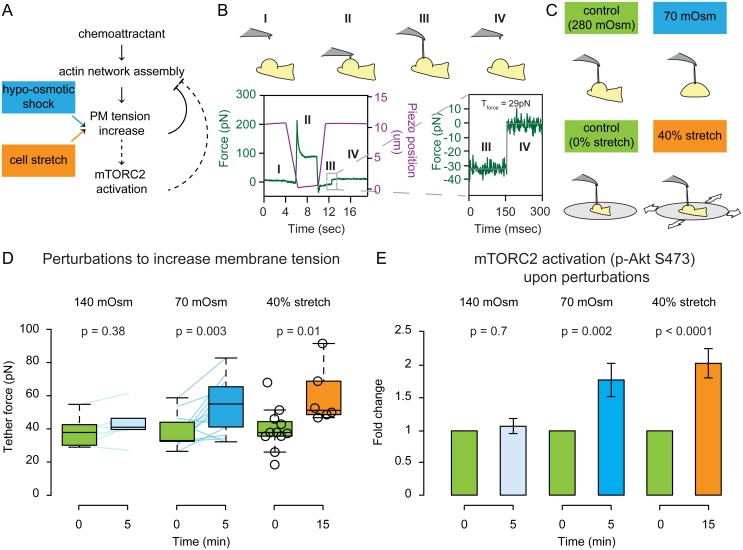
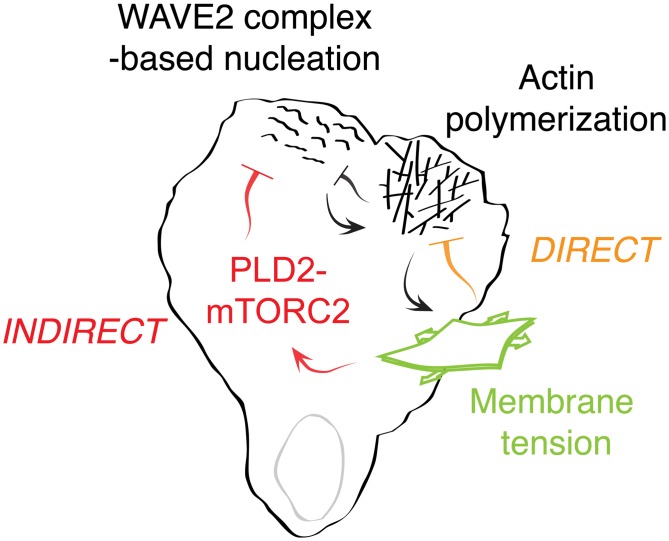
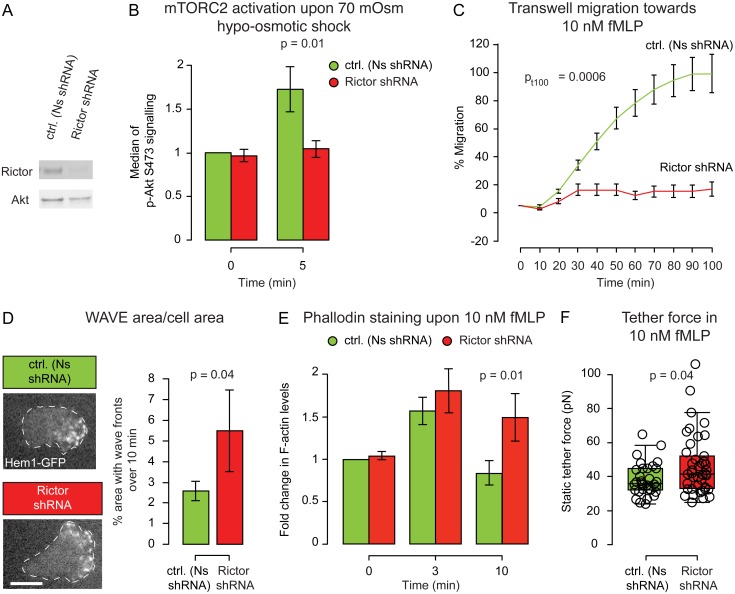
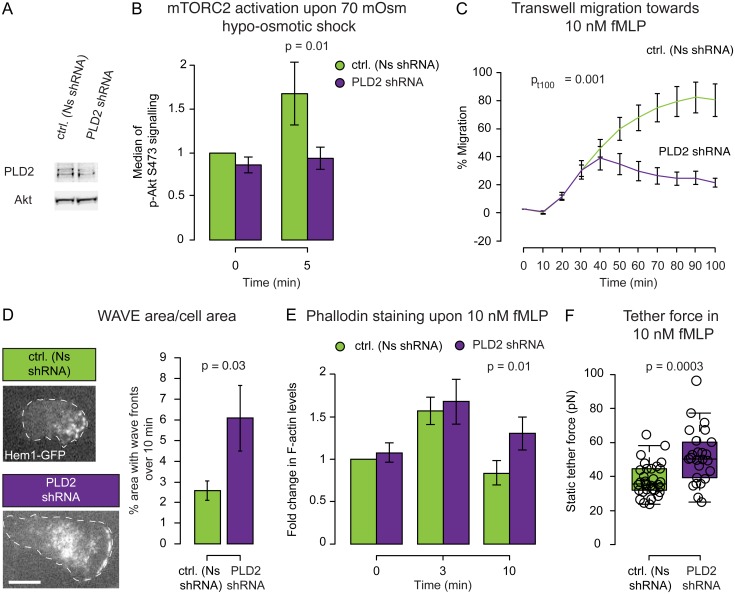
For efficient polarity and migration, cells need to regulate the magnitude and spatial distribution of actin assembly. This process is coordinated by reciprocal interactions between the actin cytoskeleton and mechanical forces. Actin polymerization-based protrusion increases tension in the plasma membrane, which in turn acts as a long-range inhibitor of actin assembly. These interactions form a negative feedback circuit that limits the magnitude of membrane tension in neutrophils and prevents expansion of the existing front and the formation of secondary fronts. It has been suggested that the plasma membrane directly inhibits actin assembly by serving as a physical barrier that opposes protrusion. Here we show that efficient control of actin polymerization-based protrusion requires an additional mechanosensory feedback cascade that indirectly links membrane tension with actin assembly. Specifically, elevated membrane tension acts through phospholipase D2 (PLD2) and the mammalian target of rapamycin complex 2 (mTORC2) to limit actin nucleation. In the absence of this pathway, neutrophils exhibit larger leading edges, higher membrane tension, and profoundly defective chemotaxis. Mathematical modeling suggests roles for both the direct (mechanical) and indirect (biochemical via PLD2 and mTORC2) feedback loops in organizing cell polarity and motility-the indirect loop is better suited to enable competition between fronts, whereas the direct loop helps spatially organize actin nucleation for efficient leading edge formation and cell movement. This circuit is essential for polarity, motility, and the control of membrane tension.
为了实现高效的极性和迁移,细胞需要调节肌动蛋白组装的程度和空间分布。这一过程由肌动蛋白细胞骨架和机械力之间的相互作用协调。基于肌动蛋白聚合的突起增加了质膜中的张力,而这种张力反过来又作为肌动蛋白组装的远程抑制剂。这些相互作用形成了一个负反馈回路,限制了中性粒细胞中膜张力的大小,并防止现有前沿的扩展和次级前沿的形成。有人提出,质膜通过作为对抗突起的物理屏障直接抑制肌动蛋白组装。在这里,我们表明,对基于肌动蛋白聚合的突起进行有效控制需要一个额外的机械传感反馈级联,该级联将膜张力与肌动蛋白组装间接联系起来。具体而言,升高的膜张力通过磷脂酶D2(PLD2)和雷帕霉素复合物2的哺乳动物靶标(mTORC2)发挥作用,以限制肌动蛋白成核。在没有这条途径的情况下,中性粒细胞表现出更大的前沿、更高的膜张力和严重缺陷的趋化性。数学建模表明,直接(机械)和间接(通过PLD2和mTORC2的生化)反馈回路在组织细胞极性和运动性方面都发挥作用——间接回路更适合于使前沿之间产生竞争,而直接回路有助于在空间上组织肌动蛋白成核,以实现高效的前沿形成和细胞运动。这个回路对于极性、运动性和膜张力的控制至关重要。