Pimentel Diogo, Donlea Jeffrey M, Talbot Clifford B, Song Seoho M, Thurston Alexander J F, Miesenböck Gero
Centre for Neural Circuits and Behaviour, University of Oxford, Tinsley Building, Mansfield Road, Oxford, OX1 3SR, United Kingdom.
Nature. 2016 Aug 18;536(7616):333-337. doi: 10.1038/nature19055. Epub 2016 Aug 3.
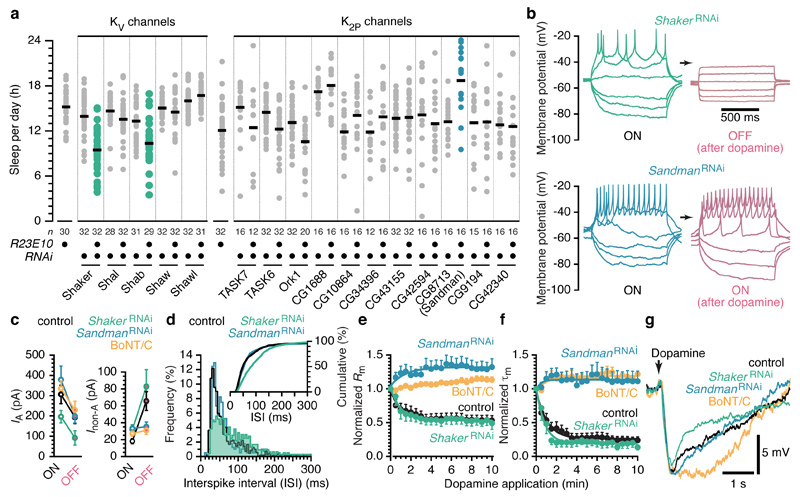
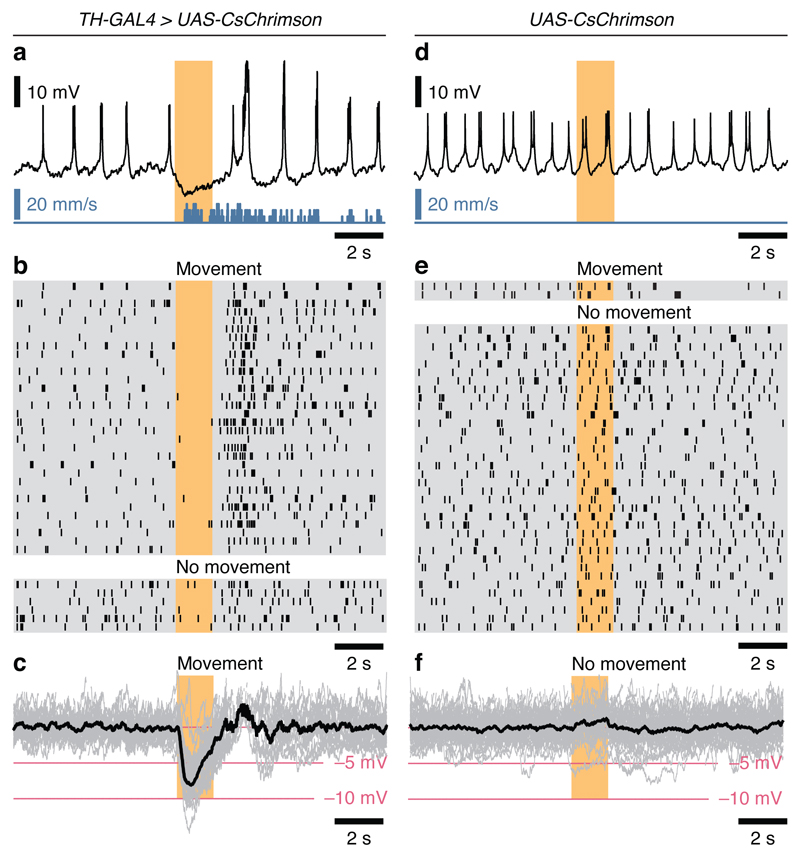
Sleep disconnects animals from the external world, at considerable risks and costs that must be offset by a vital benefit. Insight into this mysterious benefit will come from understanding sleep homeostasis: to monitor sleep need, an internal bookkeeper must track physiological changes that are linked to the core function of sleep. In Drosophila, a crucial component of the machinery for sleep homeostasis is a cluster of neurons innervating the dorsal fan-shaped body (dFB) of the central complex. Artificial activation of these cells induces sleep, whereas reductions in excitability cause insomnia. dFB neurons in sleep-deprived flies tend to be electrically active, with high input resistances and long membrane time constants, while neurons in rested flies tend to be electrically silent. Correlative evidence thus supports the simple view that homeostatic sleep control works by switching sleep-promoting neurons between active and quiescent states. Here we demonstrate state switching by dFB neurons, identify dopamine as a neuromodulator that operates the switch, and delineate the switching mechanism. Arousing dopamine caused transient hyperpolarization of dFB neurons within tens of milliseconds and lasting excitability suppression within minutes. Both effects were transduced by Dop1R2 receptors and mediated by potassium conductances. The switch to electrical silence involved the downregulation of voltage-gated A-type currents carried by Shaker and Shab, and the upregulation of voltage-independent leak currents through a two-pore-domain potassium channel that we term Sandman. Sandman is encoded by the CG8713 gene and translocates to the plasma membrane in response to dopamine. dFB-restricted interference with the expression of Shaker or Sandman decreased or increased sleep, respectively, by slowing the repetitive discharge of dFB neurons in the ON state or blocking their entry into the OFF state. Biophysical changes in a small population of neurons are thus linked to the control of sleep-wake state.
睡眠使动物与外部世界隔绝,这伴随着相当大的风险和代价,而这些必须由一项至关重要的益处来抵消。要深入了解这一神秘的益处,需从理解睡眠稳态入手:为了监测睡眠需求,一个内部的“簿记员”必须追踪与睡眠核心功能相关的生理变化。在果蝇中,睡眠稳态机制的一个关键组成部分是一群支配中央复合体背侧扇形体(dFB)的神经元。人工激活这些细胞会诱导睡眠,而兴奋性降低则会导致失眠。睡眠剥夺的果蝇中的dFB神经元往往电活动活跃,输入电阻高且膜时间常数长,而休息良好的果蝇中的神经元往往电活动静止。因此,相关证据支持了一个简单的观点,即稳态睡眠控制是通过在促进睡眠的神经元的活跃状态和静止状态之间切换来实现的。在这里,我们展示了dFB神经元的状态切换,确定多巴胺是操作该切换的神经调质,并描绘了切换机制。唤醒多巴胺会在数十毫秒内导致dFB神经元短暂超极化,并在数分钟内持续抑制兴奋性。这两种效应均由Dop1R2受体转导并由钾电导介导。向电静止状态的切换涉及由Shaker和Shab携带的电压门控A型电流的下调,以及通过我们称为Sandman的双孔域钾通道的非电压依赖性泄漏电流的上调。Sandman由CG8713基因编码,并响应多巴胺转运到质膜。对Shaker或Sandman表达的dFB特异性干扰分别通过减缓处于开启状态的dFB神经元的重复放电或阻止它们进入关闭状态来减少或增加睡眠。因此,一小群神经元的生物物理变化与睡眠 - 觉醒状态的控制相关联。