Geisler C E, Hepler C, Higgins M R, Renquist B J
School of Animal and Comparative Biomedical Sciences, University of Arizona, 4101 North Campbell Avenue, Tucson, AZ 85719 USA.
Department of Internal Medicine, University of Texas Southwestern Medical Center, Dallas, TX 75235 USA.
Nutr Metab (Lond). 2016 Sep 26;13:62. doi: 10.1186/s12986-016-0122-x. eCollection 2016.
The increased incidence of obesity and associated metabolic diseases has driven research focused on genetically or pharmacologically alleviating metabolic dysfunction. These studies employ a range of fasting-refeeding models including 4-24 h fasts, "overnight" fasts, or meal feeding. Still, we lack literature that describes the physiologically relevant adaptations that accompany changes in the duration of fasting and re-feeding. Since the liver is central to whole body metabolic homeostasis, we investigated the timing of the fast-induced shift toward glycogenolysis, gluconeogenesis, and ketogenesis and the meal-induced switch toward glycogenesis and away from ketogenesis.
Twelve to fourteen week old male C57BL/6J mice were fasted for 0, 4, 8, 12, or 16 h and sacrificed 4 h after lights on. In a second study, designed to understand the response to a meal, we gave fasted mice access to feed for 1 or 2 h before sacrifice. We analyzed the data using mixed model analysis of variance.
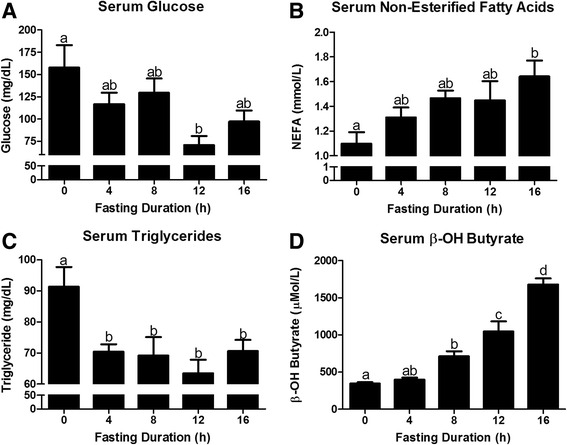
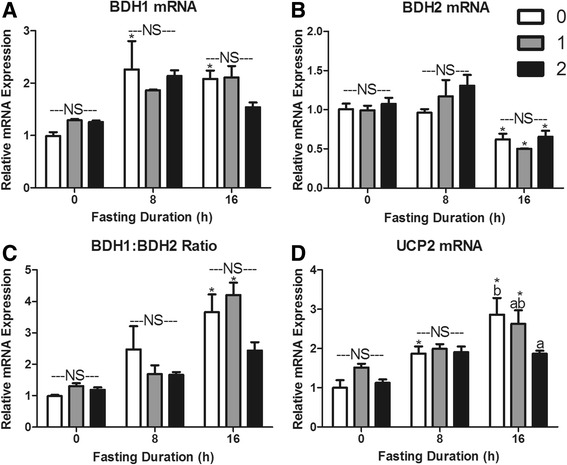
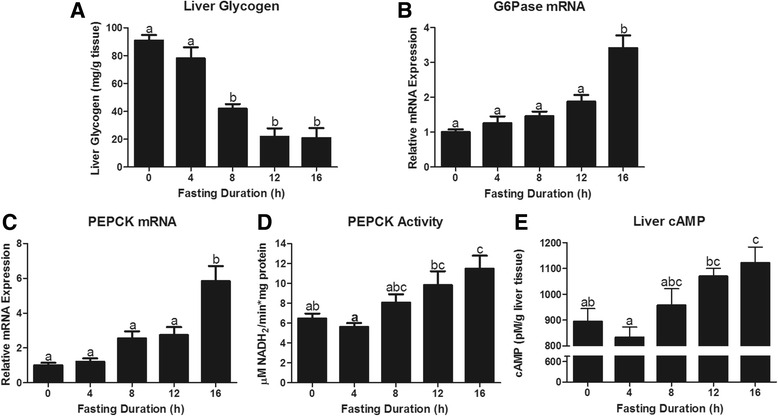
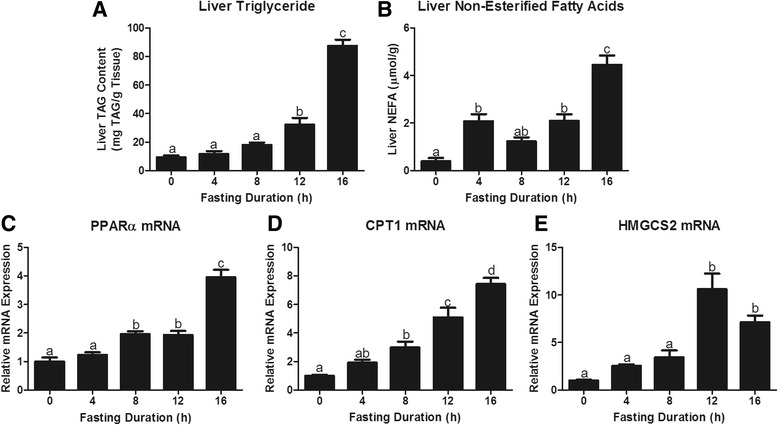
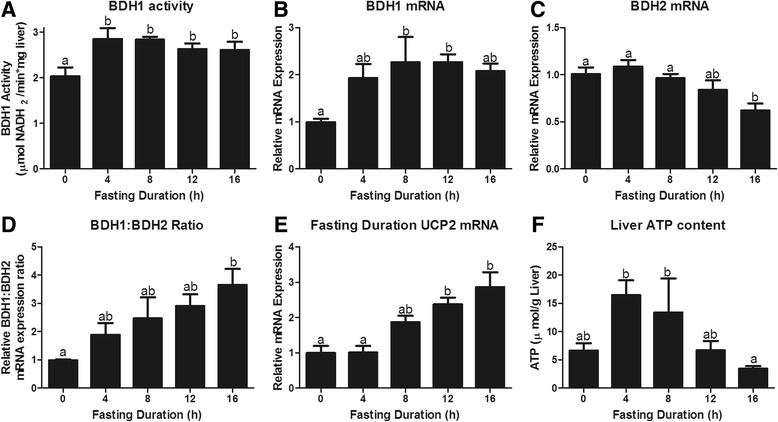
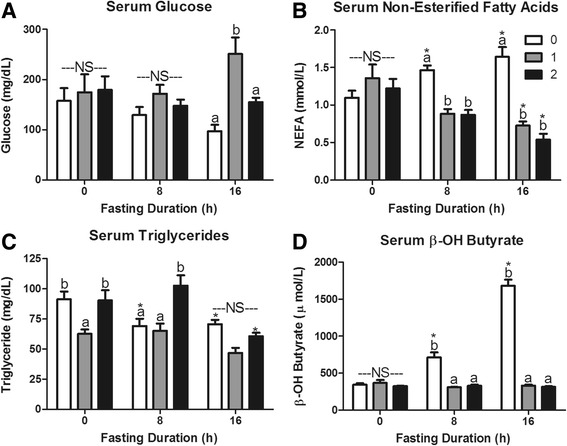
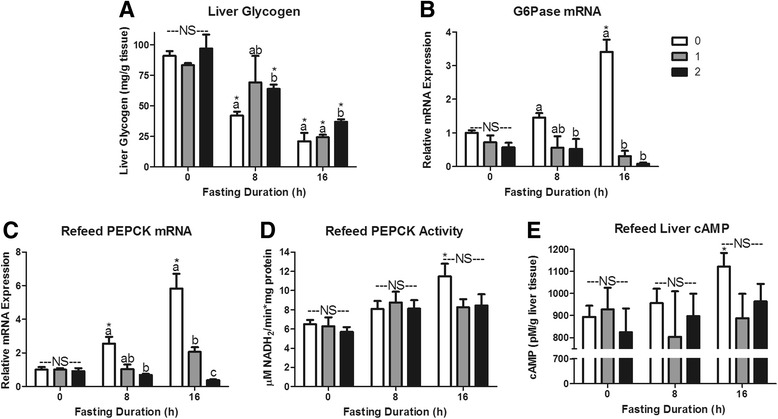
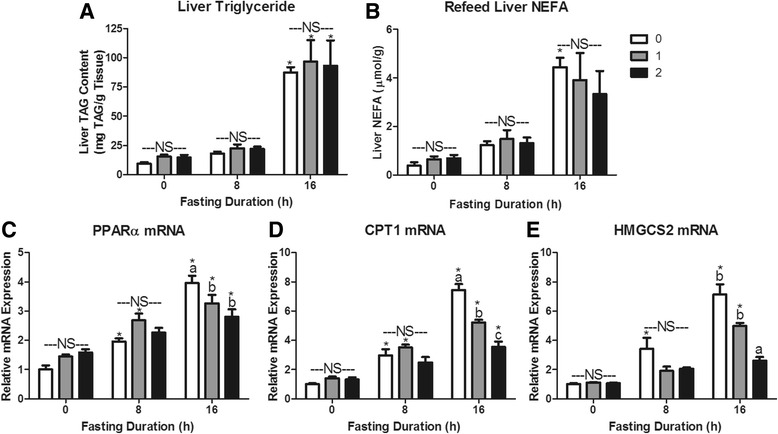
Fasting initiated robust metabolic shifts, evidenced by changes in serum glucose, non-esterified fatty acids (NEFAs), triacylglycerol, and β-OH butyrate, as well as, liver triacylglycerol, non-esterified fatty acid, and glycogen content. Glycogenolysis is the primary source to maintain serum glucose during the first 8 h of fasting, while de novo gluconeogenesis is the primary source thereafter. The increase in serum β-OH butyrate results from increased enzymatic capacity for fatty acid flux through β-oxidation and shunting of acetyl-CoA toward ketone body synthesis (increased CPT1 (Carnitine Palmitoyltransferase 1) and HMGCS2 (3-Hydroxy-3-Methylglutaryl-CoA Synthase 2) expression, respectively). In opposition to the relatively slow metabolic adaptation to fasting, feeding of a meal results in rapid metabolic changes including full depression of serum β-OH butyrate and NEFAs within an hour.
Herein, we provide a detailed description of timing of the metabolic adaptations in response to fasting and re-feeding to inform study design in experiments of metabolic homeostasis. Since fasting and obesity are both characterized by elevated adipose tissue lipolysis, hepatic lipid accumulation, ketogenesis, and gluconeogenesis, understanding the drivers behind the metabolic shift from the fasted to the fed state may provide targets to limit aberrant gluconeogenesis and ketogenesis in obesity.
肥胖及相关代谢性疾病发病率的上升推动了旨在通过基因或药物手段缓解代谢功能障碍的研究。这些研究采用了一系列禁食-再进食模型,包括4 - 24小时禁食、“过夜”禁食或进食。然而,我们缺乏描述禁食和再进食持续时间变化所伴随的生理相关适应性变化的文献。由于肝脏在全身代谢稳态中起核心作用,我们研究了禁食诱导的向糖原分解、糖异生和酮体生成转变的时间,以及进食诱导的向糖原合成转变和远离酮体生成的时间。
将12至14周龄的雄性C57BL/6J小鼠禁食0、4、8、12或16小时,并在光照后4小时处死。在第二项旨在了解对进食反应的研究中,我们在处死前让禁食的小鼠进食1或2小时。我们使用方差混合模型分析来分析数据。
禁食引发了显著的代谢变化,血清葡萄糖、非酯化脂肪酸(NEFAs)、三酰甘油和β-羟基丁酸的变化以及肝脏三酰甘油、非酯化脂肪酸和糖原含量的变化证明了这一点。糖原分解是禁食最初8小时内维持血清葡萄糖的主要来源,而此后从头开始的糖异生是主要来源。血清β-羟基丁酸的增加源于通过β-氧化的脂肪酸通量的酶能力增加以及乙酰辅酶A向酮体合成的分流(分别是肉碱棕榈酰转移酶1(CPT1)和3-羟基-3-甲基戊二酰辅酶A合酶2(HMGCS2)表达增加)。与对禁食相对缓慢的代谢适应相反,进食一餐会导致快速的代谢变化,包括一小时内血清β-羟基丁酸和NEFAs完全降低。
在此,我们详细描述了对禁食和再进食的代谢适应时间,为代谢稳态实验的研究设计提供信息。由于禁食和肥胖都以脂肪组织脂解增加、肝脏脂质积累、酮体生成和糖异生为特征,了解从禁食状态到进食状态代谢转变背后的驱动因素可能为限制肥胖中异常的糖异生和酮体生成提供靶点。