Goldstein Ido, Baek Songjoon, Presman Diego M, Paakinaho Ville, Swinstead Erin E, Hager Gordon L
Laboratory of Receptor Biology and Gene Expression, The National Cancer Institute, The National Institutes of Health, Bethesda, Maryland 20892, USA.
Genome Res. 2017 Mar;27(3):427-439. doi: 10.1101/gr.212175.116. Epub 2016 Dec 28.
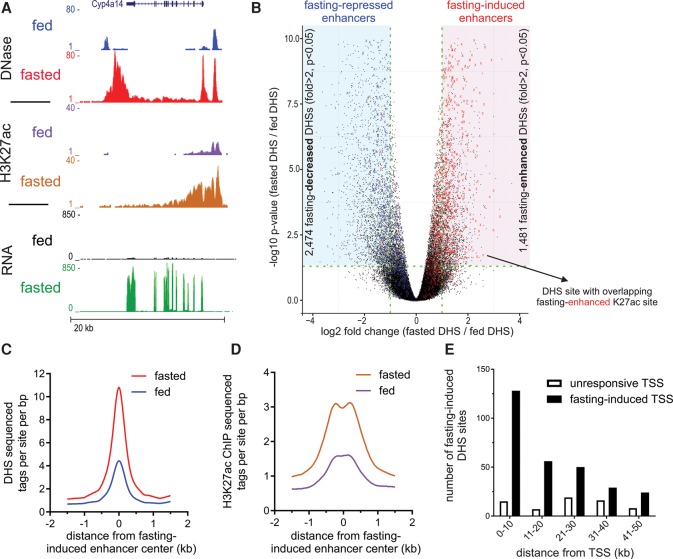
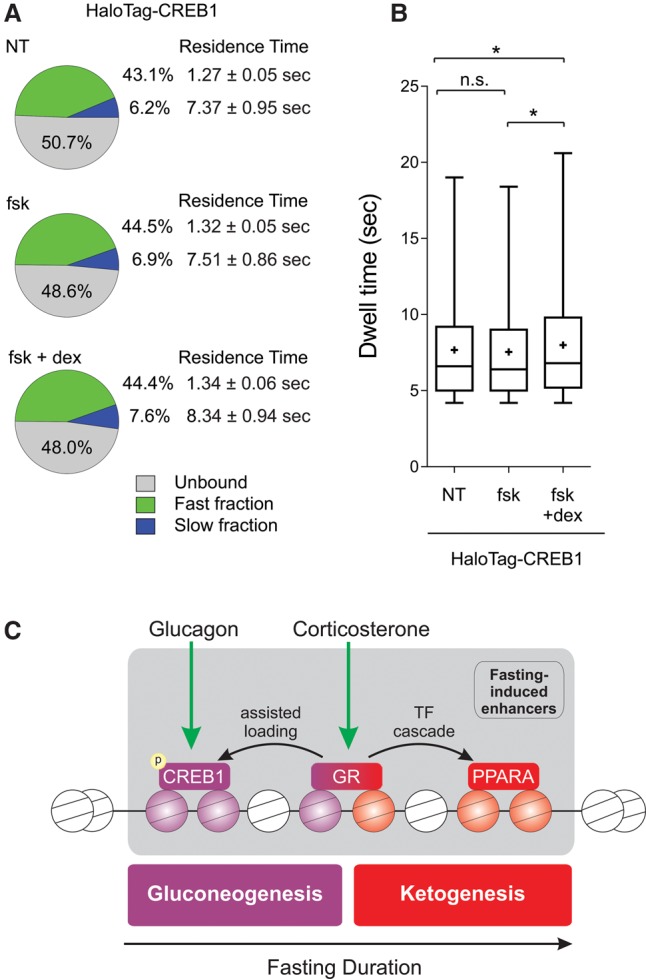
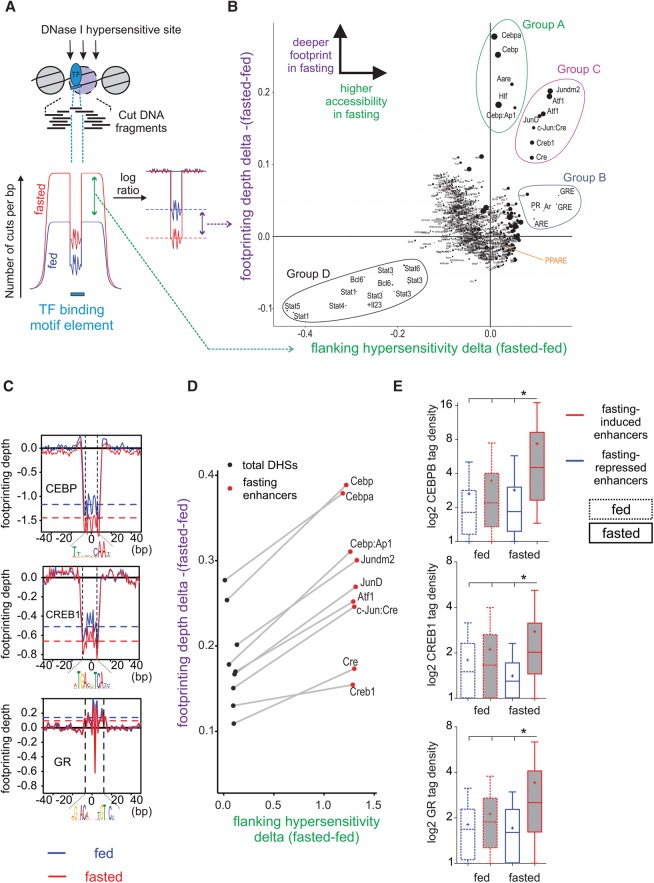
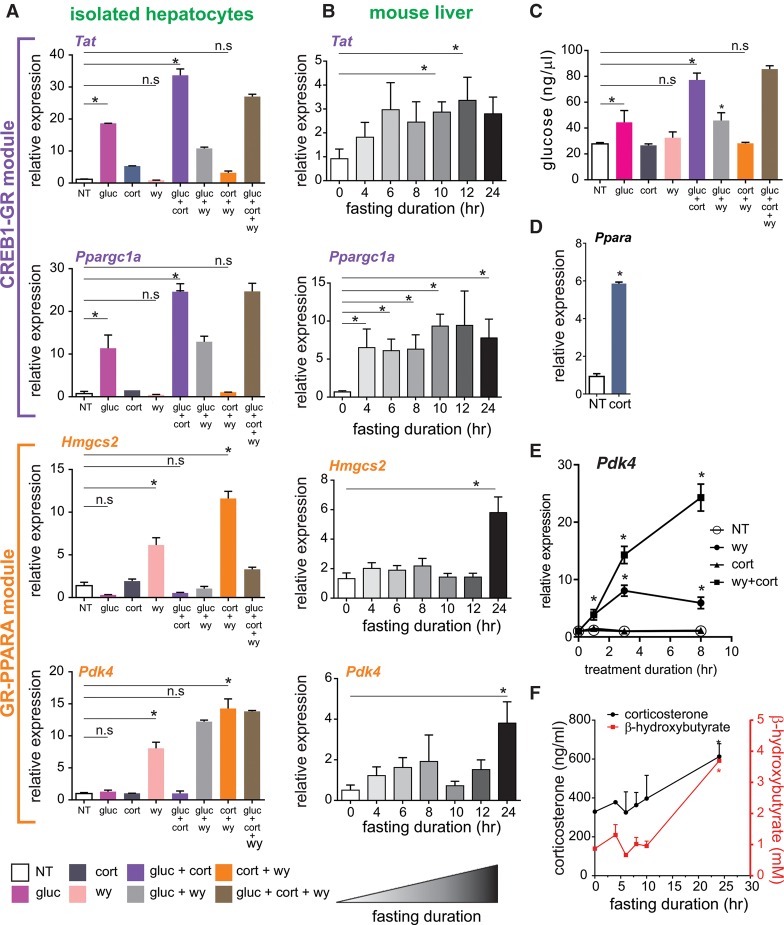
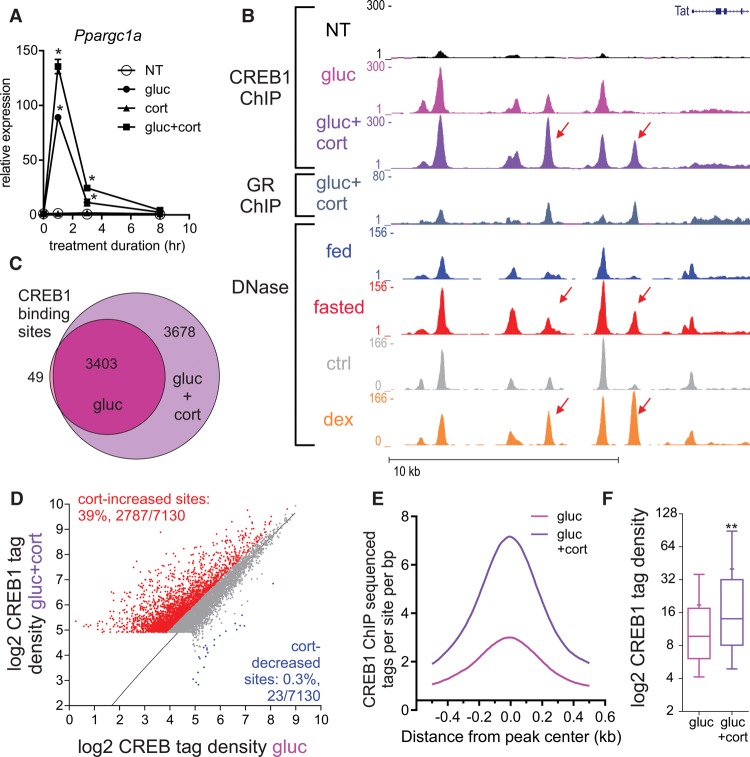
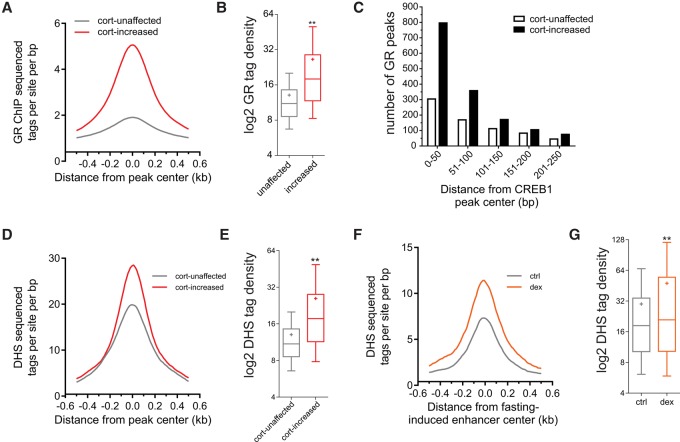
Fasting elicits transcriptional programs in hepatocytes leading to glucose and ketone production. This transcriptional program is regulated by many transcription factors (TFs). To understand how this complex network regulates the metabolic response to fasting, we aimed at isolating the enhancers and TFs dictating it. Measuring chromatin accessibility revealed that fasting massively reorganizes liver chromatin, exposing numerous fasting-induced enhancers. By utilizing computational methods in combination with dissecting enhancer features and TF cistromes, we implicated four key TFs regulating the fasting response: glucocorticoid receptor (GR), cAMP responsive element binding protein 1 (CREB1), peroxisome proliferator activated receptor alpha (PPARA), and CCAAT/enhancer binding protein beta (CEBPB). These TFs regulate fuel production by two distinctly operating modules, each controlling a separate metabolic pathway. The gluconeogenic module operates through assisted loading, whereby GR doubles the number of sites occupied by CREB1 as well as enhances CREB1 binding intensity and increases accessibility of CREB1 binding sites. Importantly, this GR-assisted CREB1 binding was enhancer-selective and did not affect all CREB1-bound enhancers. Single-molecule tracking revealed that GR increases the number and DNA residence time of a portion of chromatin-bound CREB1 molecules. These events collectively result in rapid synergistic gene expression and higher hepatic glucose production. Conversely, the ketogenic module operates via a GR-induced TF cascade, whereby PPARA levels are increased following GR activation, facilitating gradual enhancer maturation next to PPARA target genes and delayed ketogenic gene expression. Our findings reveal a complex network of enhancers and TFs that dynamically cooperate to restore homeostasis upon fasting.
禁食会引发肝细胞中的转录程序,导致葡萄糖和酮的生成。这个转录程序受许多转录因子(TFs)调控。为了理解这个复杂的网络如何调节对禁食的代谢反应,我们旨在分离决定该反应的增强子和转录因子。测量染色质可及性发现,禁食会大规模重组肝脏染色质,暴露出大量禁食诱导的增强子。通过结合计算方法以及剖析增强子特征和转录因子顺式作用元件组,我们确定了四个调节禁食反应的关键转录因子:糖皮质激素受体(GR)、环磷酸腺苷反应元件结合蛋白1(CREB1)、过氧化物酶体增殖物激活受体α(PPARA)和CCAAT/增强子结合蛋白β(CEBPB)。这些转录因子通过两个截然不同的运作模块调节燃料生成,每个模块控制一条独立的代谢途径。糖异生模块通过辅助加载起作用,即GR使CREB1占据的位点数量翻倍,并增强CREB1的结合强度,同时增加CREB1结合位点的可及性。重要的是,这种GR辅助的CREB1结合具有增强子选择性,并不影响所有与CREB1结合的增强子。单分子追踪显示,GR增加了一部分与染色质结合的CREB1分子的数量和在DNA上的停留时间。这些事件共同导致快速的协同基因表达和更高的肝脏葡萄糖生成。相反,生酮模块通过GR诱导的转录因子级联起作用,即GR激活后PPARA水平升高,促进PPARA靶基因附近增强子的逐渐成熟以及延迟生酮基因的表达。我们的研究结果揭示了一个由增强子和转录因子组成的复杂网络,它们在禁食时动态协作以恢复体内平衡。