Mendoza-Becerril María A, Marian José Eduardo A R, Migotto Alvaro Esteves, Marques Antonio Carlos
Department of Zoology, Institute of Biosciences, University of São Paulo , São Paulo , Brazil.
Department of Zoology, Institute of Biosciences, University of São Paulo, São Paulo, Brazil; Center for Marine Biology, University of São Paulo, São Sebastião, São Paulo, Brazil.
PeerJ. 2017 Feb 16;5:e2964. doi: 10.7717/peerj.2964. eCollection 2017.
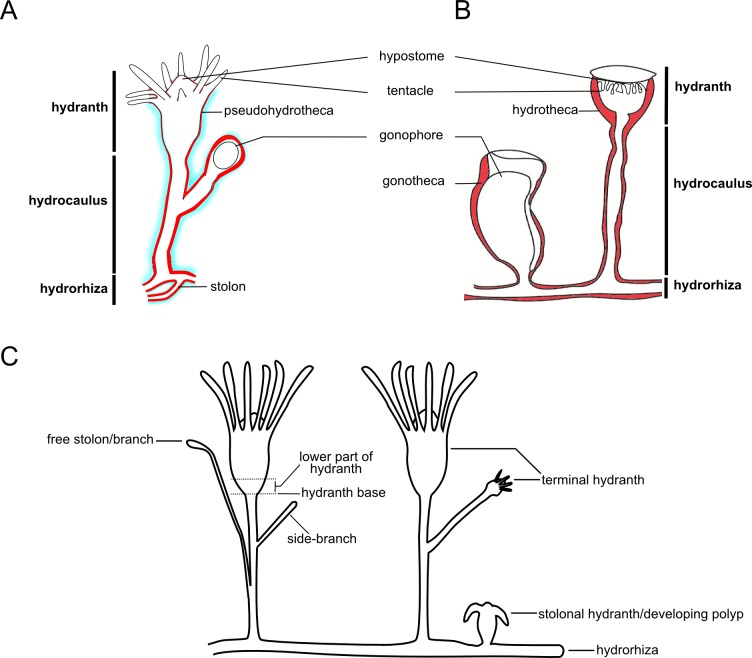
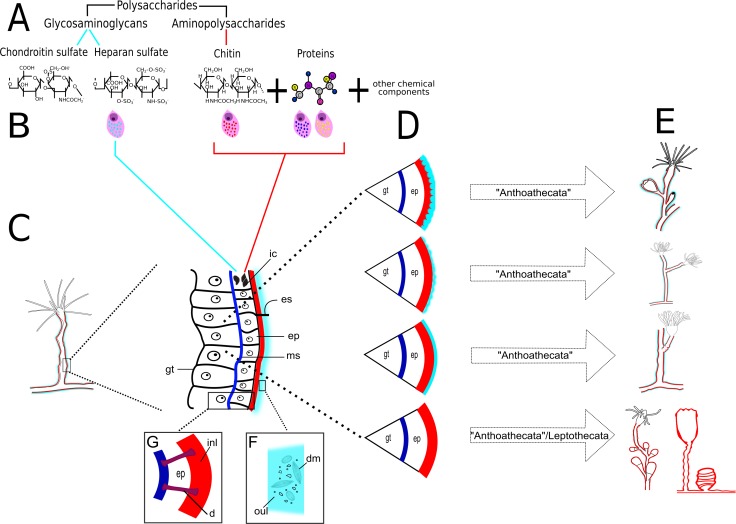
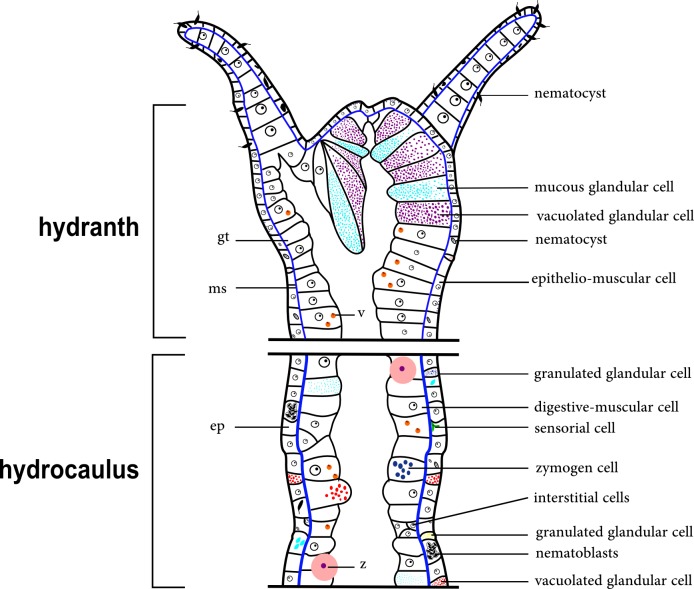
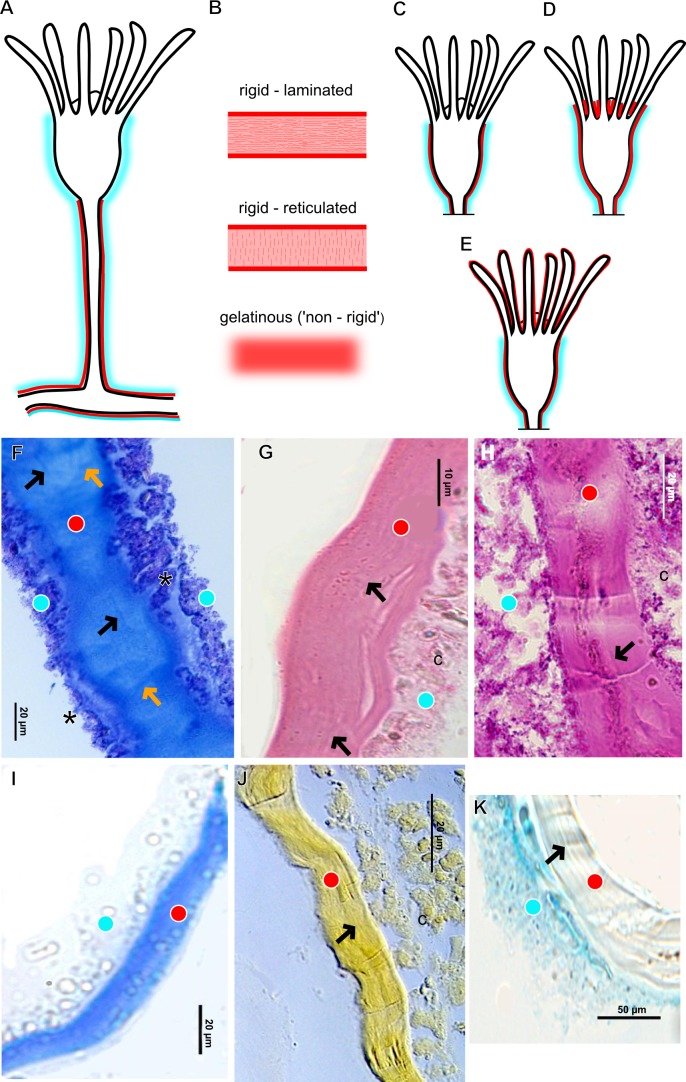
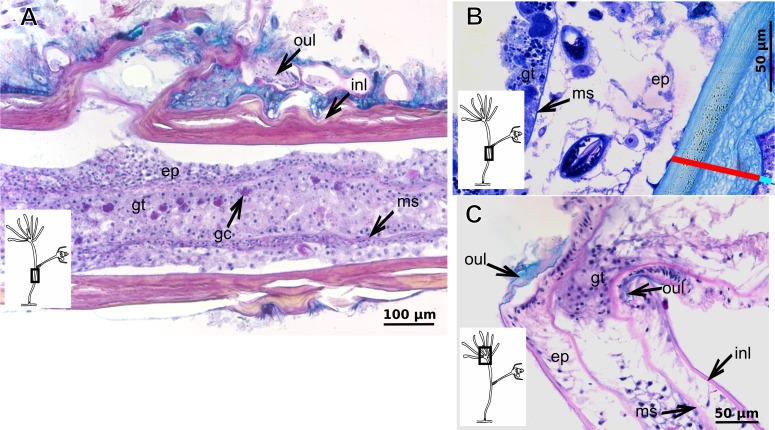
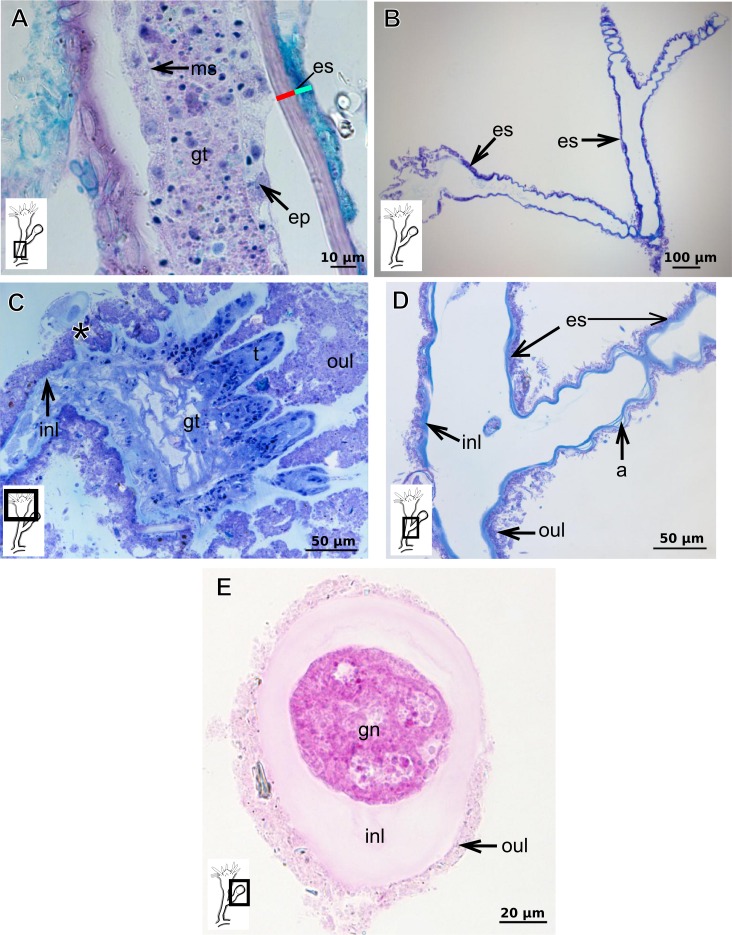
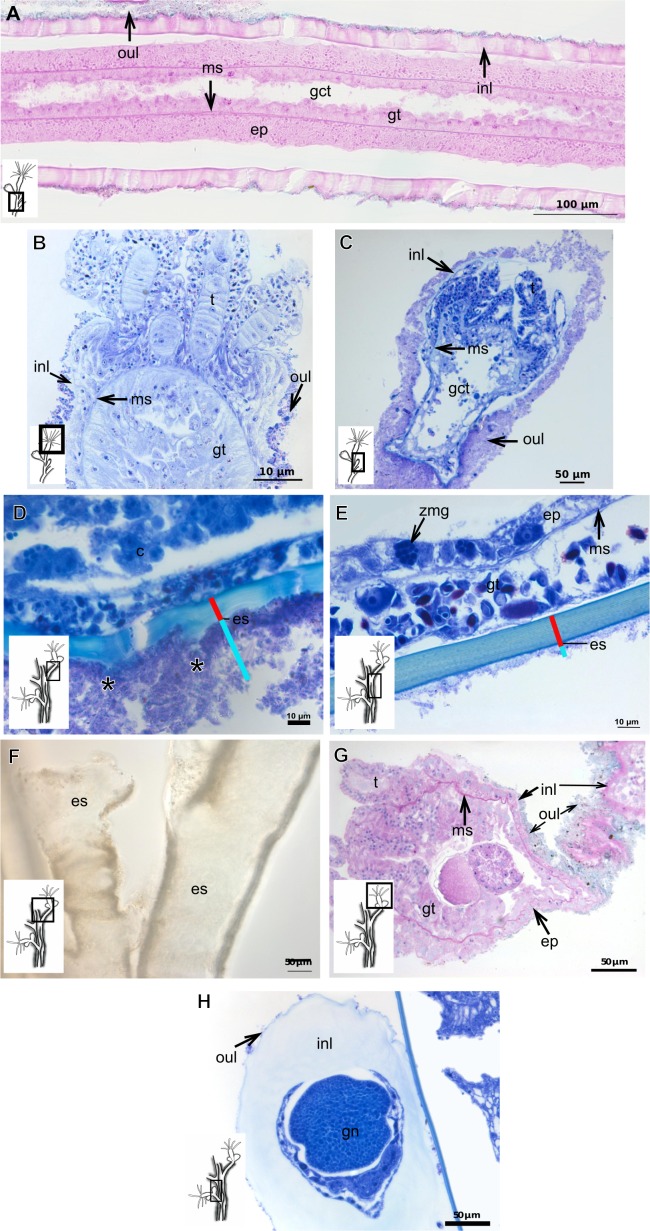
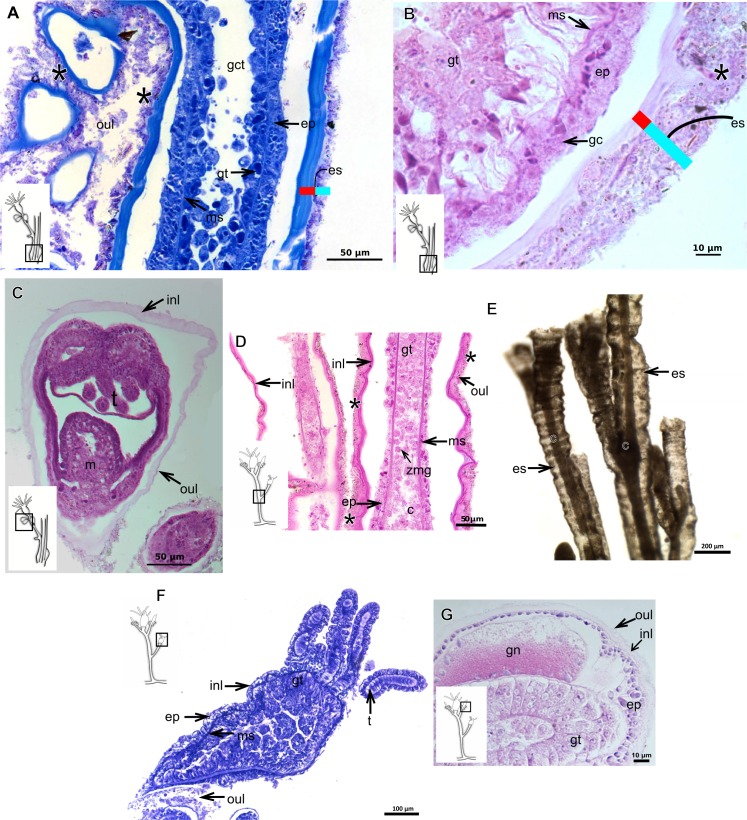
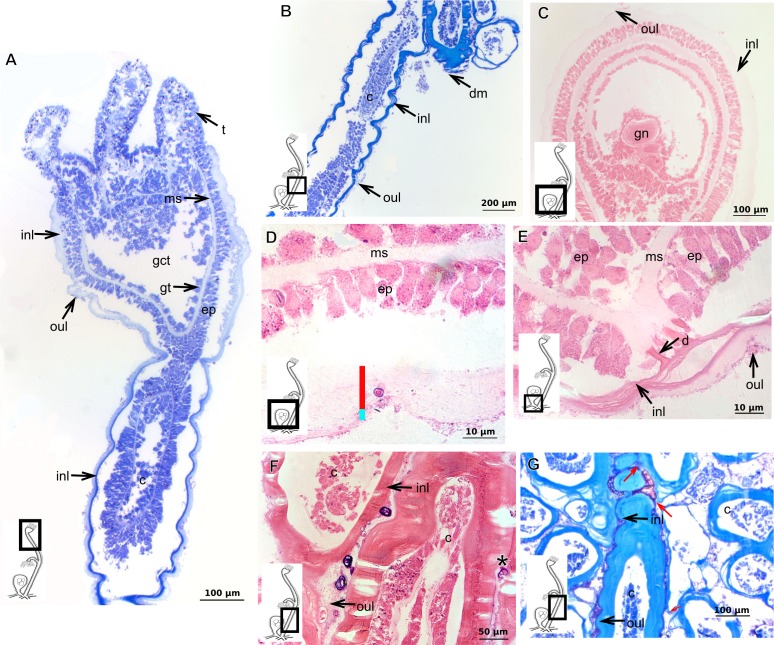
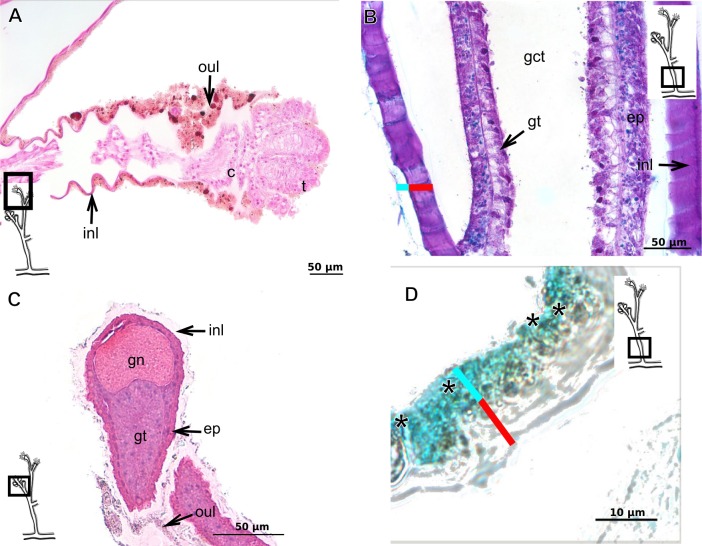
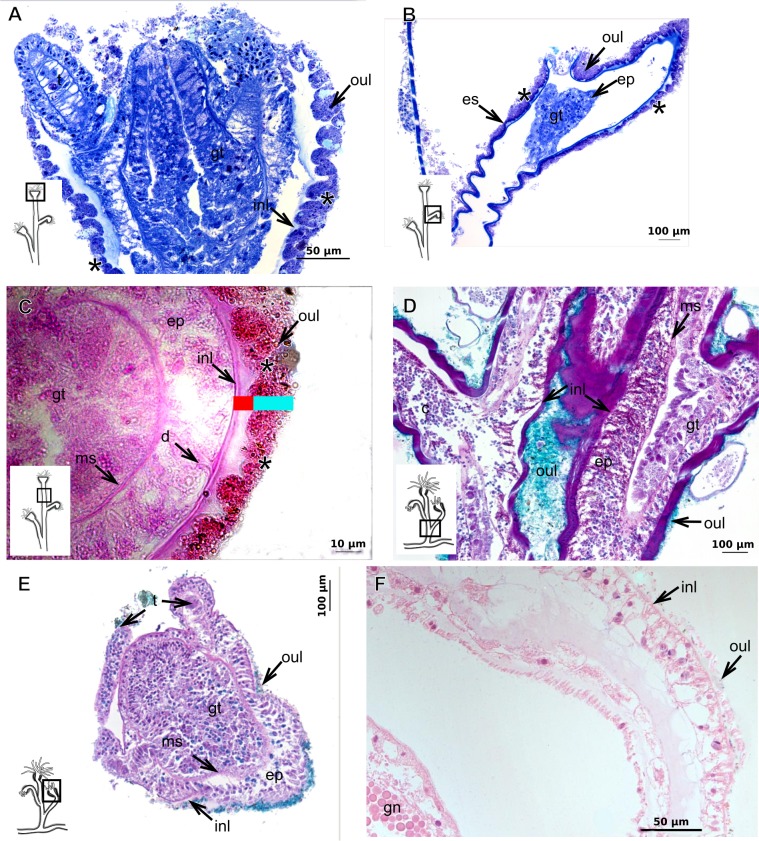
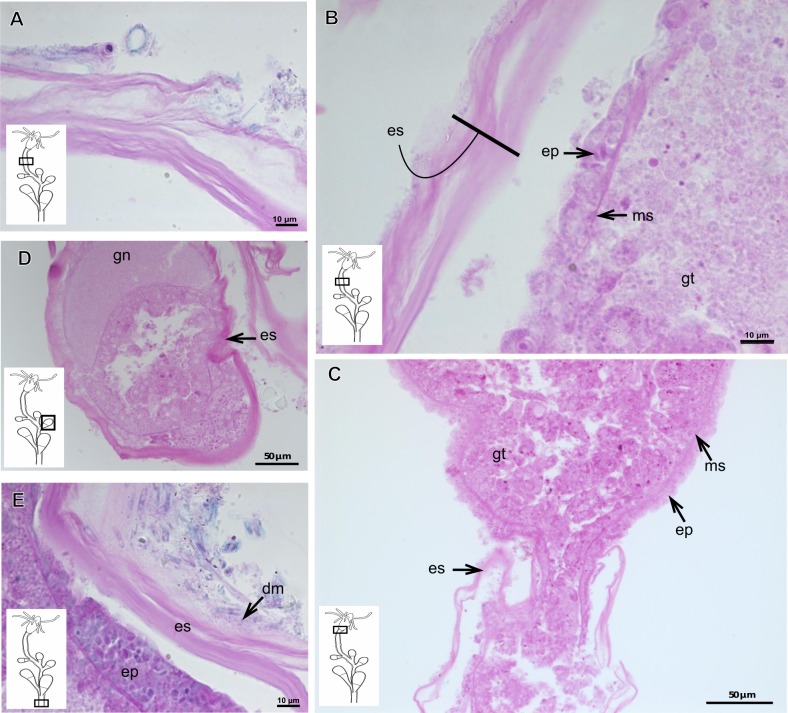
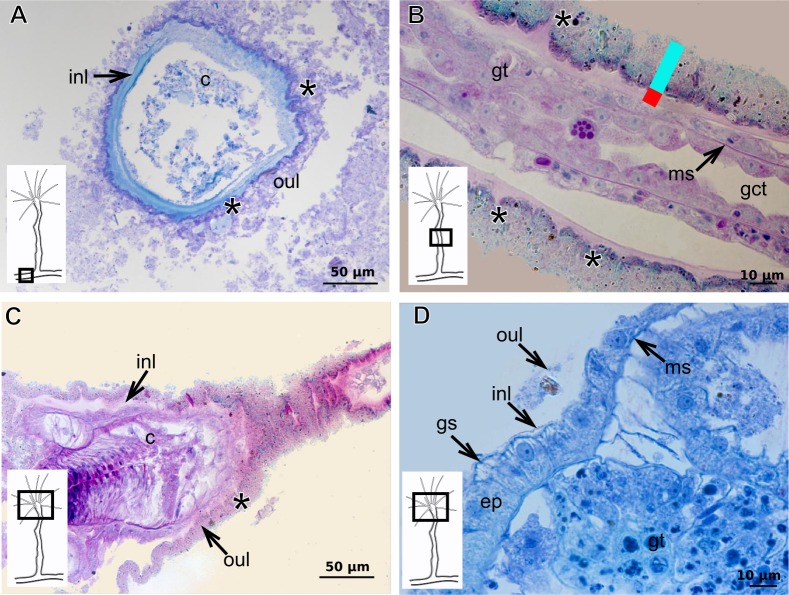
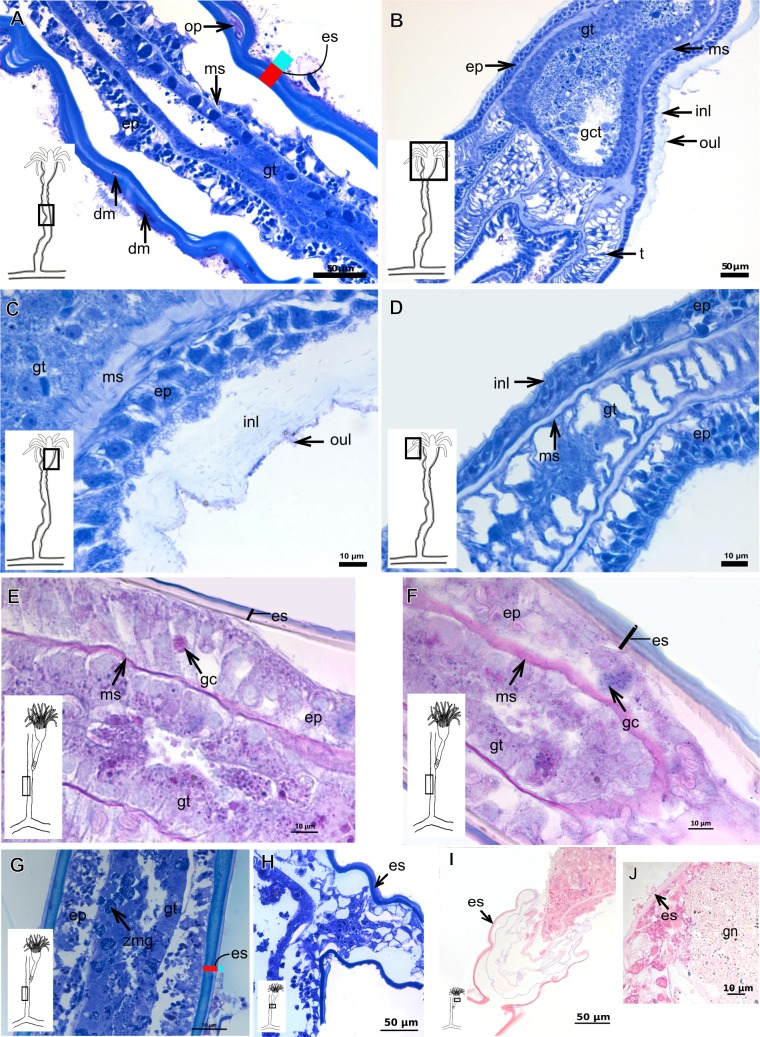
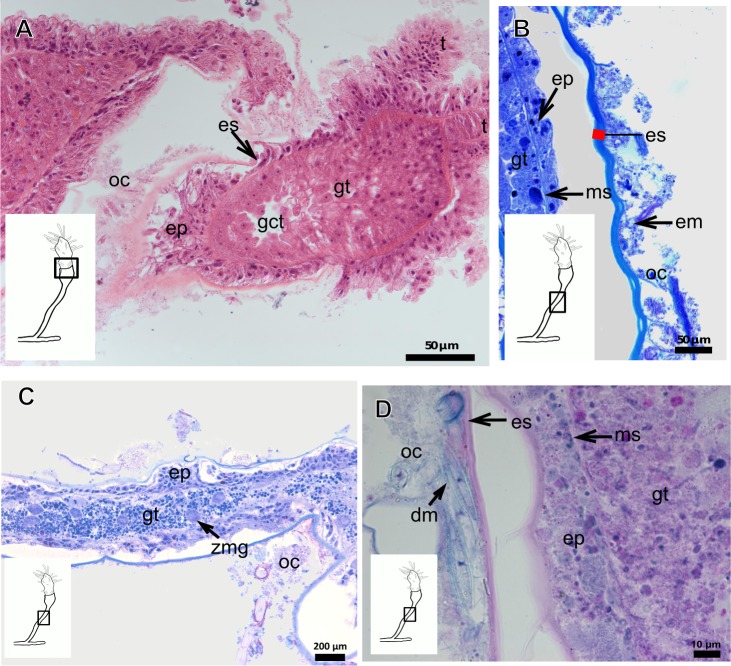
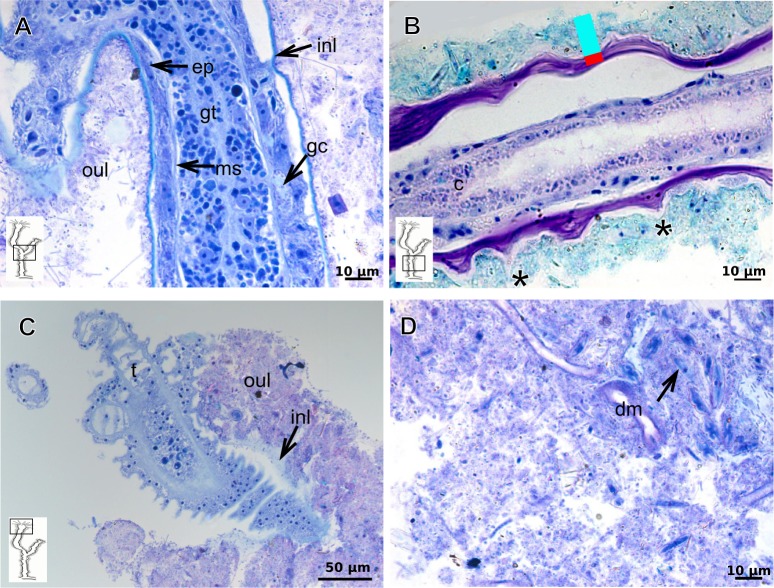
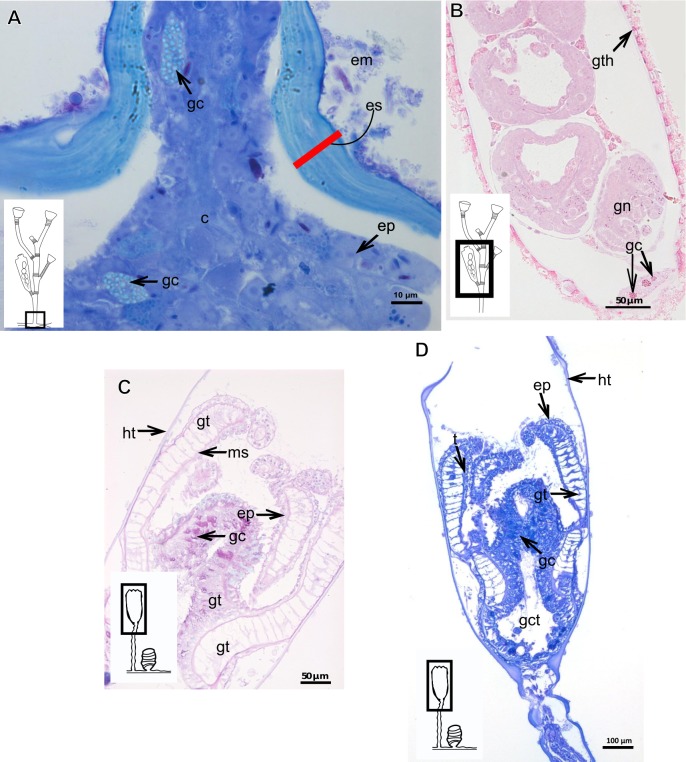
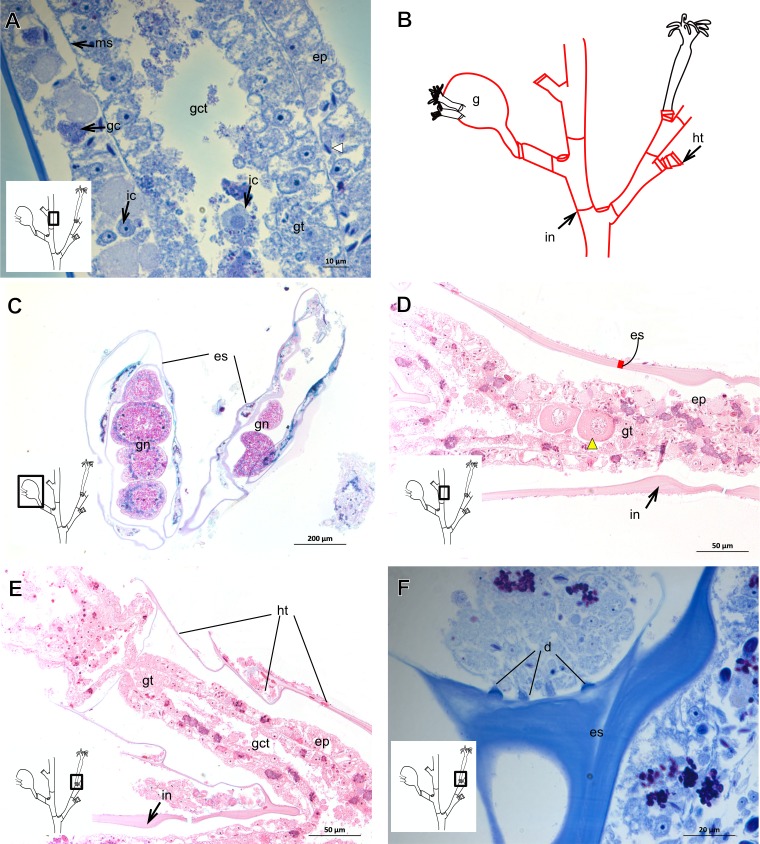
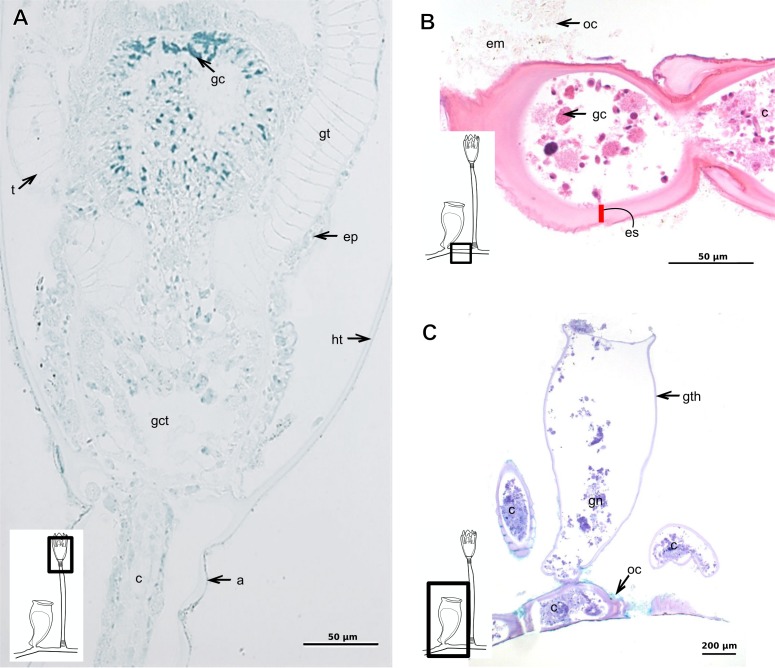
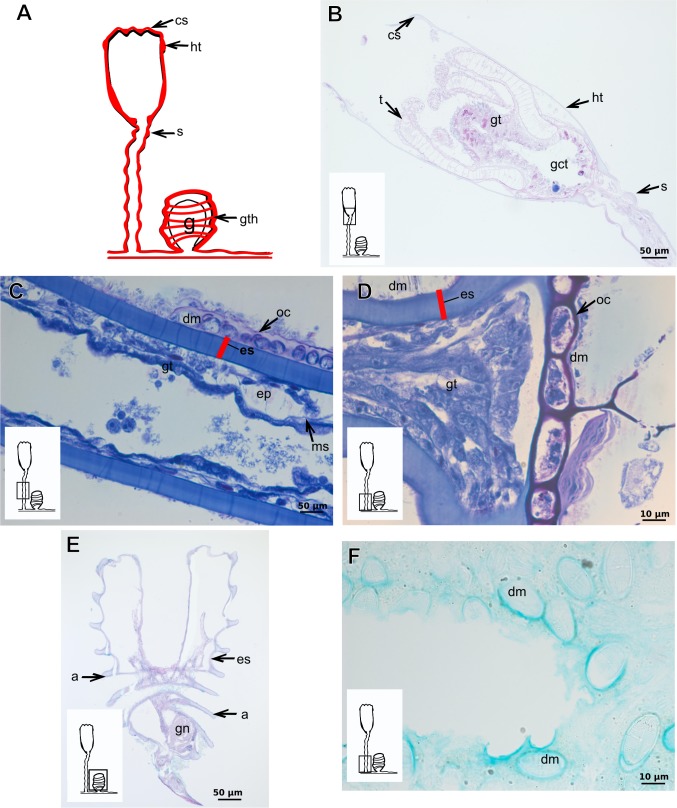
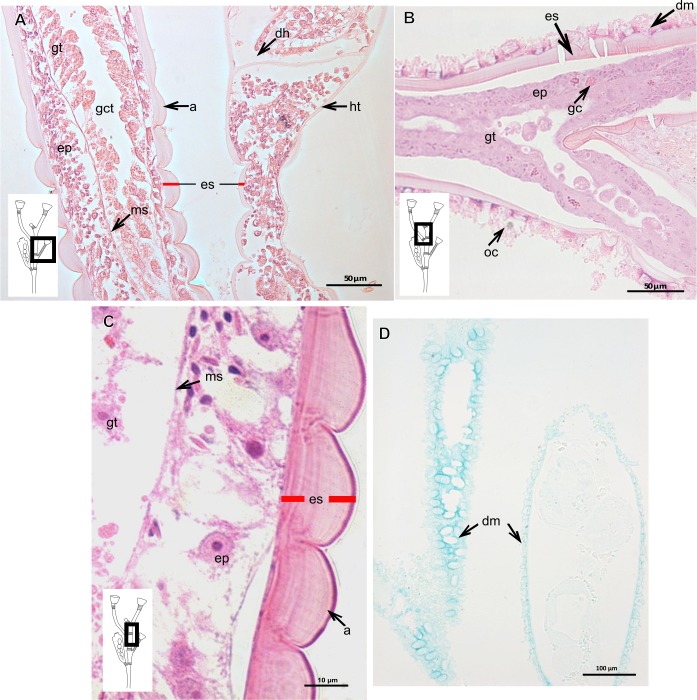
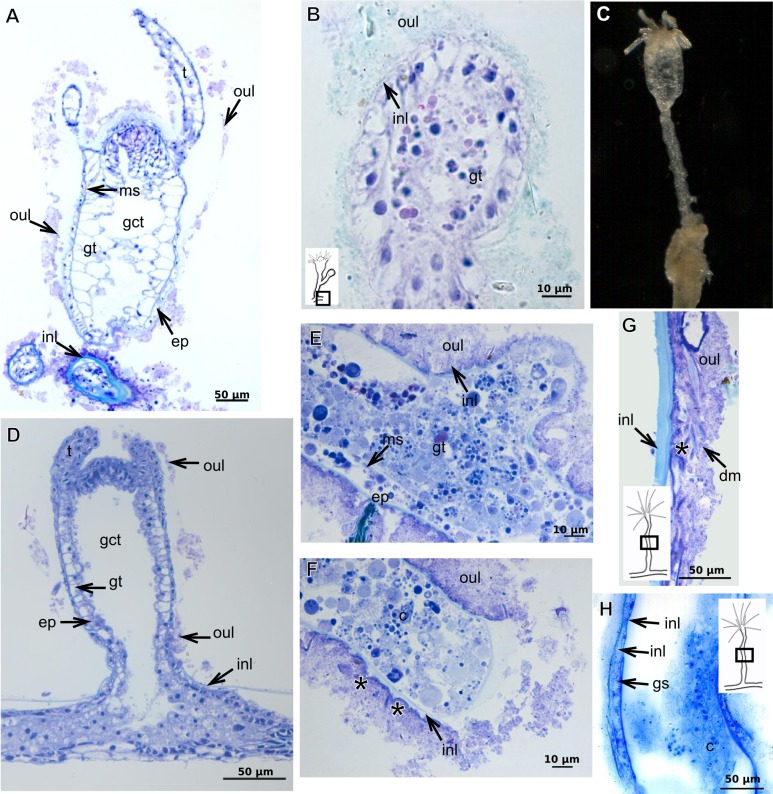
The exoskeleton is an important source of characters for the taxonomy of Hydroidolina. It originates as epidermal secretions and, among other functions, protects the coenosarc of the polypoid stage. However, comparative studies on the exoskeletal tissue origin, development, chemical, and structural characteristics, as well as its evolution and homology, are few and fragmented. This study compares the structure and composition of the exoskeleton and underlying coenosarc in members of "Anthoathecata" and some Leptothecata, but does so mainly in bougainvilliid polyps histological analyses. We also studied the development of the exoskeleton under experimental conditions. We identified three types of glandular epidermal cells related to the origin of the exoskeleton and the secretion of its polysaccharides component. The exoskeleton of the species studied is either bilayered (perisarc and exosarc, especially in bougainvilliids) or corneous (perisarc). The exoskeleton varies in chemical composition, structural rigidity, thickness, extension, and coverage in the different regions of the colony. In bilayered exoskeletons, the exosarc is produced first and appears to be a key step in the formation of the rigid exoskeleton. The exoskeleton contains anchoring structures such as desmocytes and "perisarc extensions."
外骨骼是水螅虫纲分类学中重要的特征来源。它起源于表皮分泌物,除其他功能外,还保护着螅形体阶段的共肉。然而,关于外骨骼组织的起源、发育、化学和结构特征,以及其进化和同源性的比较研究很少且零散。本研究比较了“花水母目”和一些软水母目的成员中外骨骼和其下方共肉的结构与组成,但主要是在布氏螅体的组织学分析中进行的。我们还在实验条件下研究了外骨骼的发育。我们确定了三种与外骨骼起源及其多糖成分分泌相关的腺状表皮细胞。所研究物种的外骨骼要么是双层的(围鞘和外鞘,特别是在布氏螅中),要么是角质的(围鞘)。外骨骼在群体的不同区域,其化学成分、结构硬度、厚度、延伸范围和覆盖程度各不相同。在双层外骨骼中,外鞘首先产生,似乎是形成坚硬外骨骼的关键步骤。外骨骼包含诸如连细胞和“围鞘延伸物”等锚定结构。