Magaraki Aristea, van der Heijden Godfried, Sleddens-Linkels Esther, Magarakis Leonidas, van Cappellen Wiggert A, Peters Antoine H F M, Gribnau Joost, Baarends Willy M, Eijpe Maureen
Department of Developmental Biology, Erasmus MC, University Medical Center, Rotterdam, The Netherlands.
Division of Reproductive Medicine, Department of Obstetrics and Gynecology, Erasmus MC, Rotterdam, The Netherlands.
Epigenetics Chromatin. 2017 Mar 11;10:11. doi: 10.1186/s13072-017-0119-3. eCollection 2017.
In the nuclei of most mammalian cells, pericentric heterochromatin is characterized by DNA methylation, histone modifications such as H3K9me3 and H4K20me3, and specific binding proteins like heterochromatin-binding protein 1 isoforms (HP1 isoforms). Maintenance of this specialized chromatin structure is of great importance for genome integrity and for the controlled repression of the repetitive elements within the pericentric DNA sequence. Here we have studied histone modifications at pericentric heterochromatin during primordial germ cell (PGC) development using different fixation conditions and fluorescent immunohistochemical and immunocytochemical protocols.
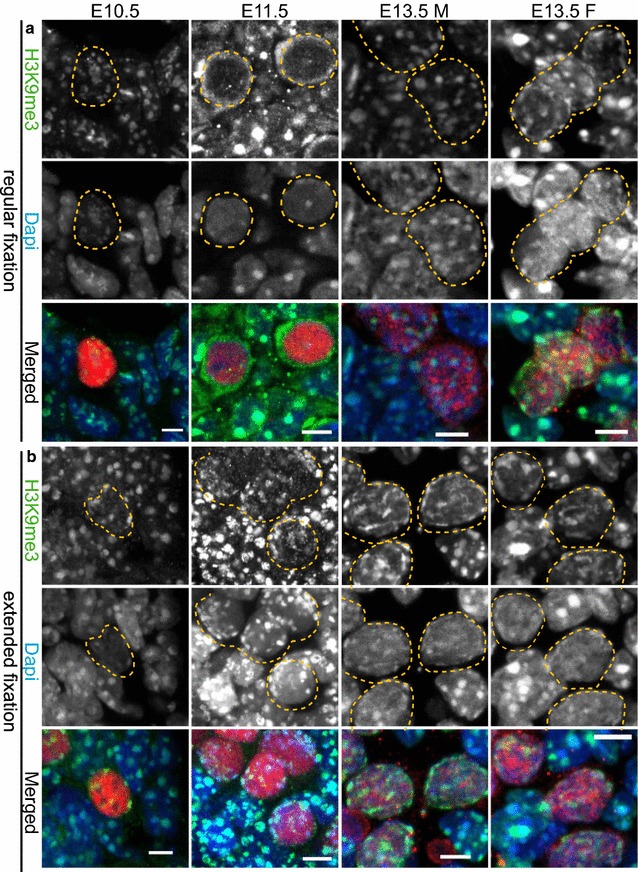
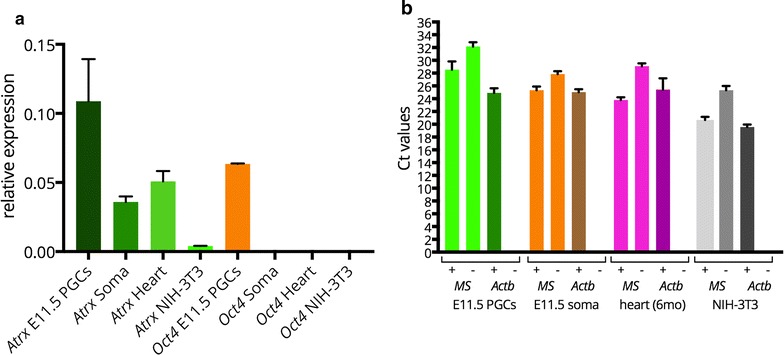
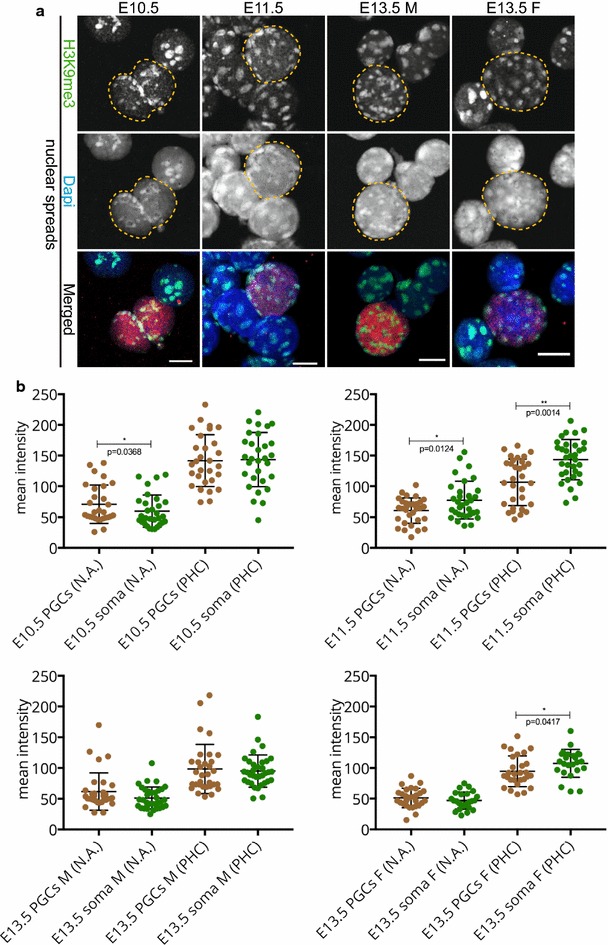
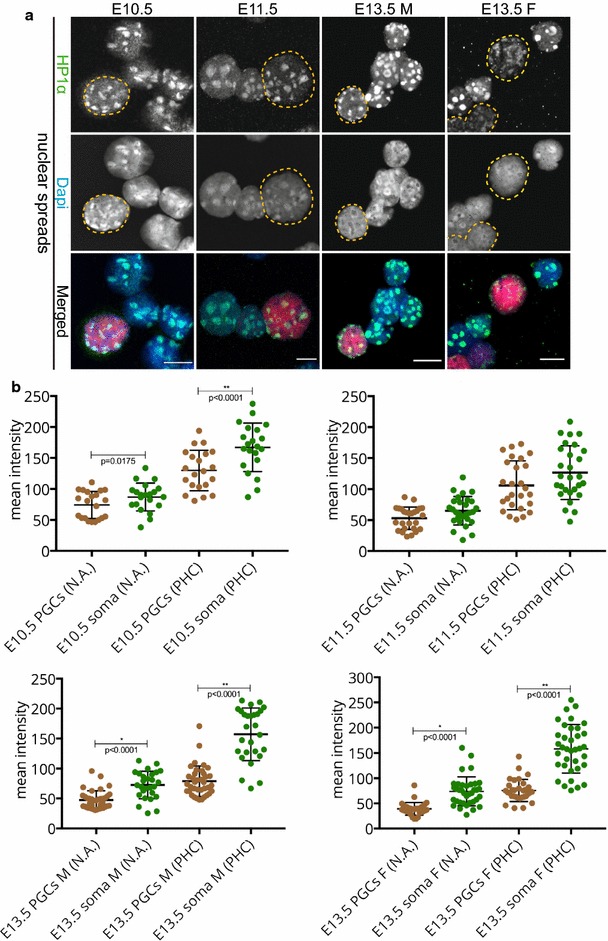
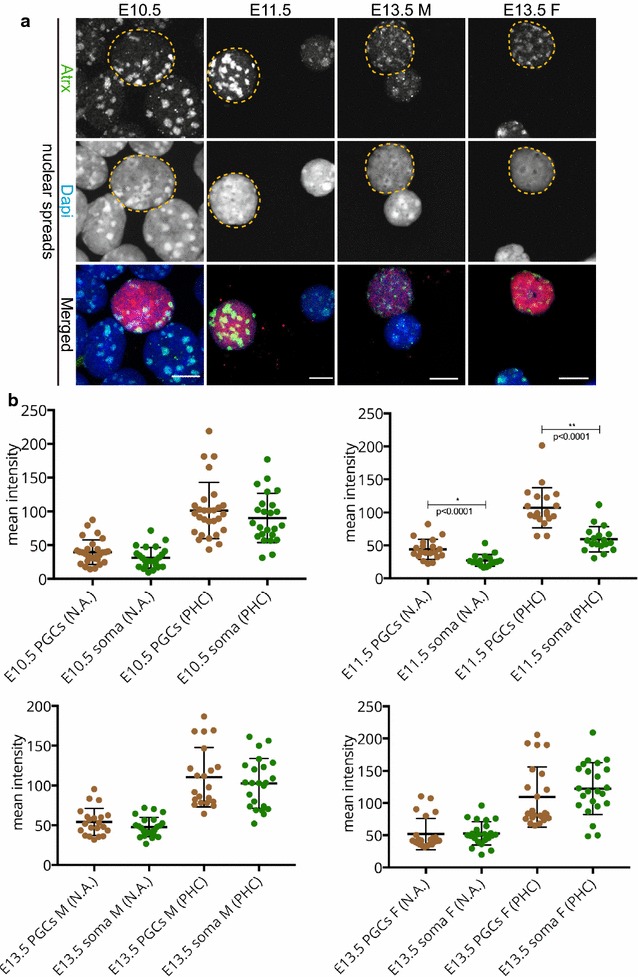
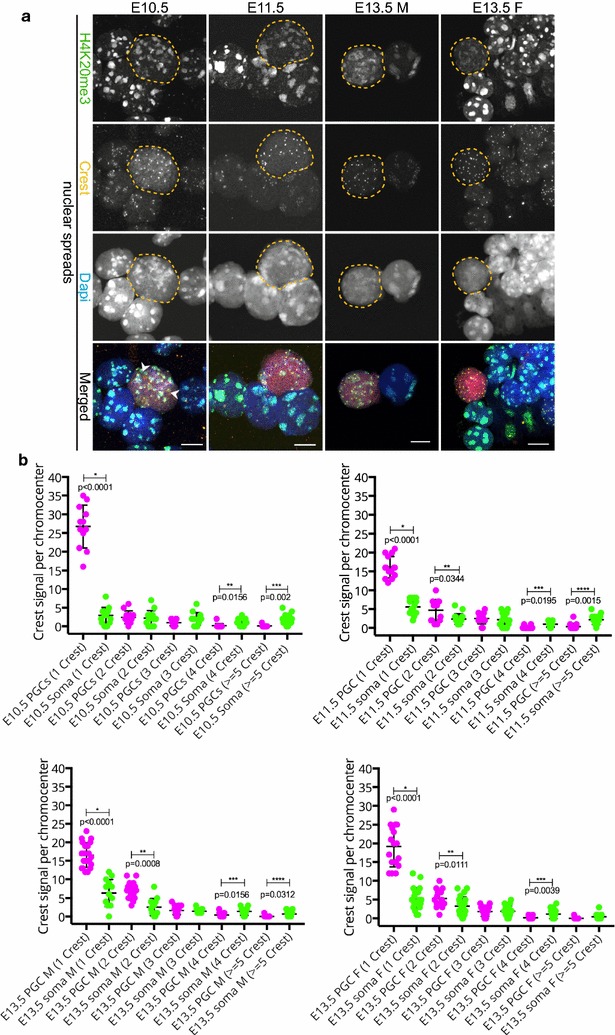
We observed that pericentric heterochromatin marks, such as H3K9me3, H4K20me3, and HP1 isoforms, were retained on pericentric heterochromatin throughout PGC development. However, the observed immunostaining patterns varied, depending on the fixation method, explaining previous findings of a general loss of pericentric heterochromatic features in PGCs. Also, in contrast to the general clustering of multiple pericentric regions and associated centromeres in DAPI-dense regions in somatic cells, the pericentric regions of PGCs were more frequently organized as individual entities. We also observed a transient enrichment of the chromatin remodeler ATRX in pericentric regions in embryonic day 11.5 (E11.5) PGCs. At this stage, a similar and low level of major satellite repeat RNA transcription was detected in both PGCs and somatic cells.
These results indicate that in pericentric heterochromatin of mouse PGCs, only minor reductions in levels of some chromatin-associated proteins occur, in association with a transient increase in ATRX, between E11.5 and E13.5. These pericentric heterochromatin regions more frequently contain only a single centromere in PGCs compared to the surrounding soma, indicating a difference in overall organization, but there is no de-repression of major satellite transcription.
在大多数哺乳动物细胞的细胞核中,着丝粒周围异染色质的特征包括DNA甲基化、组蛋白修饰(如H3K9me3和H4K20me3)以及特定结合蛋白(如异染色质结合蛋白1亚型(HP1亚型))。维持这种特殊的染色质结构对于基因组完整性以及着丝粒周围DNA序列中重复元件的受控抑制至关重要。在此,我们使用不同的固定条件以及荧光免疫组织化学和免疫细胞化学方法,研究了原始生殖细胞(PGC)发育过程中着丝粒周围异染色质的组蛋白修饰。
我们观察到,着丝粒周围异染色质标记,如H3K9me3、H4K20me3和HP1亚型,在整个PGC发育过程中都保留在着丝粒周围异染色质上。然而,观察到的免疫染色模式因固定方法而异,这解释了之前关于PGC中着丝粒周围异染色质特征普遍丧失的研究结果。此外,与体细胞中多个着丝粒周围区域和相关着丝粒在DAPI致密区域的一般聚集情况不同,PGC的着丝粒周围区域更常作为单个实体组织起来。我们还观察到,在胚胎第11.5天(E11.5)的PGC中,染色质重塑因子ATRX在着丝粒周围区域有短暂富集。在此阶段,在PGC和体细胞中均检测到相似且低水平的主要卫星重复RNA转录。
这些结果表明,在小鼠PGC的着丝粒周围异染色质中,在E11.5和E13.5之间,某些染色质相关蛋白水平仅略有降低,同时伴有ATRX的短暂增加。与周围体细胞相比,这些着丝粒周围异染色质区域在PGC中更常仅包含单个着丝粒,这表明整体组织存在差异,但主要卫星转录并未去抑制。