Fukasawa Yoshinori, Oda Toshiyuki, Tomii Kentaro, Imai Kenichiro
Artificial Intelligence Research Center, National Institute of Advanced Science and Technology (AIST), Tokyo, Japan.
Biotechnology Research Institute for Drug Discovery, National Institute of Advanced Science and Technology (AIST), Tokyo, Japan.
Mol Biol Evol. 2017 Jul 1;34(7):1574-1586. doi: 10.1093/molbev/msx096.
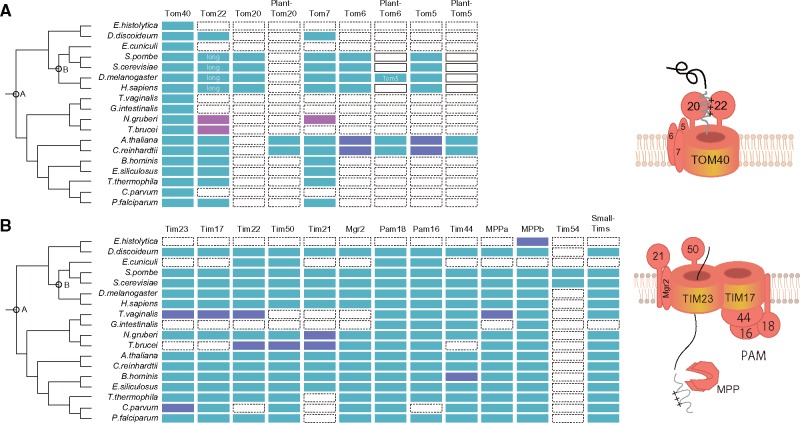
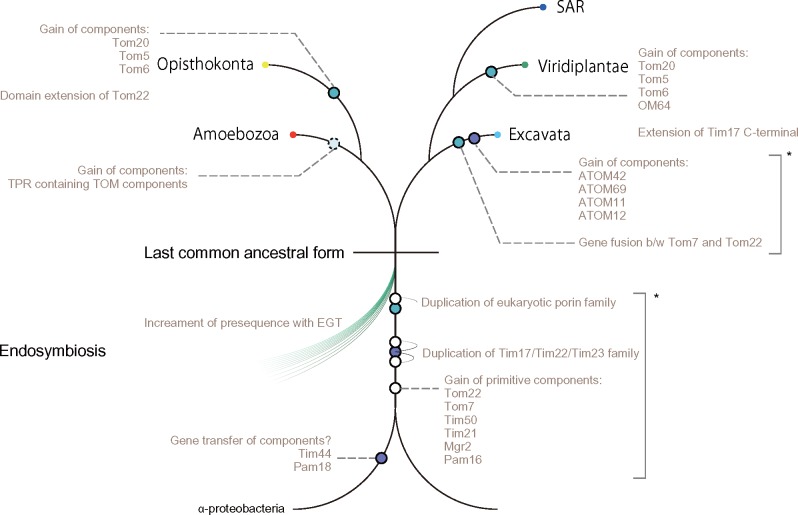
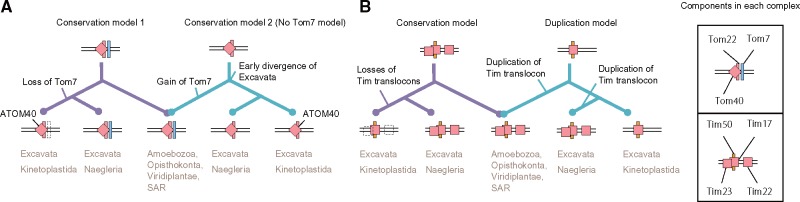
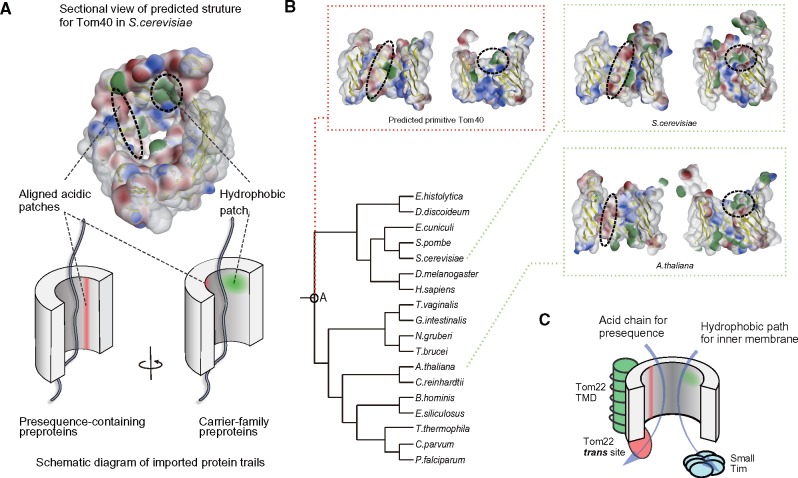
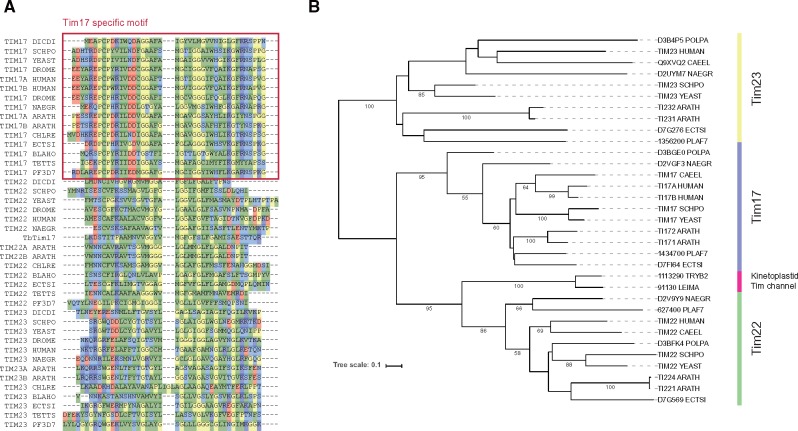
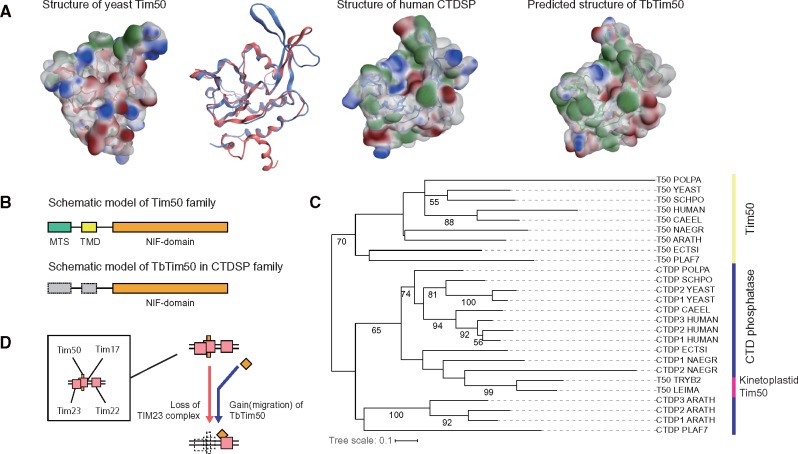
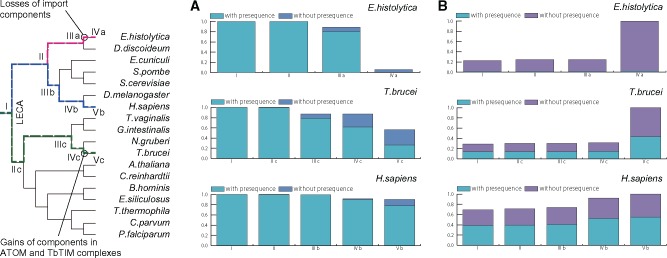
Protein transport systems are fundamentally important for maintaining mitochondrial function. Nevertheless, mitochondrial protein translocases such as the kinetoplastid ATOM complex have recently been shown to vary in eukaryotic lineages. Various evolutionary hypotheses have been formulated to explain this diversity. To resolve any contradiction, estimating the primitive state and clarifying changes from that state are necessary. Here, we present more likely primitive models of mitochondrial translocases, specifically the translocase of the outer membrane (TOM) and translocase of the inner membrane (TIM) complexes, using scrutinized phylogenetic profiles. We then analyzed the translocases' evolution in eukaryotic lineages. Based on those results, we propose a novel evolutionary scenario for diversification of the mitochondrial transport system. Our results indicate that presequence transport machinery was mostly established in the last eukaryotic common ancestor, and that primitive translocases already had a pathway for transporting presequence-containing proteins. Moreover, secondary changes including convergent and migrational gains of a presequence receptor in TOM and TIM complexes, respectively, likely resulted from constrained evolution. The nature of a targeting signal can constrain alteration to the protein transport complex.
蛋白质转运系统对于维持线粒体功能至关重要。然而,近来发现诸如动质体ATOM复合物等线粒体蛋白质转运酶在真核生物谱系中存在差异。人们已经提出了各种进化假说以解释这种多样性。为了解决任何矛盾之处,估计原始状态并阐明从该状态发生的变化是必要的。在此,我们利用仔细审查的系统发育概况,提出了更可能的线粒体转运酶原始模型,特别是外膜转运酶(TOM)和内膜转运酶(TIM)复合物。然后,我们分析了真核生物谱系中线粒体转运酶的进化。基于这些结果,我们提出了线粒体运输系统多样化的一种新的进化情景。我们的结果表明,前序列转运机制大多是在最后的真核生物共同祖先中建立起来的,并且原始转运酶已经具有一条用于运输含前序列蛋白质的途径。此外,包括分别在TOM和TIM复合物中前序列受体的趋同和迁移性获得等二级变化,可能是由受限进化导致的。靶向信号的性质会限制蛋白质转运复合物的改变。