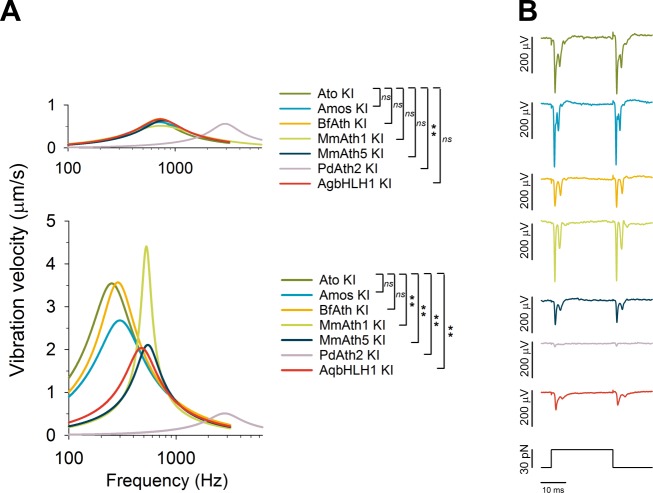
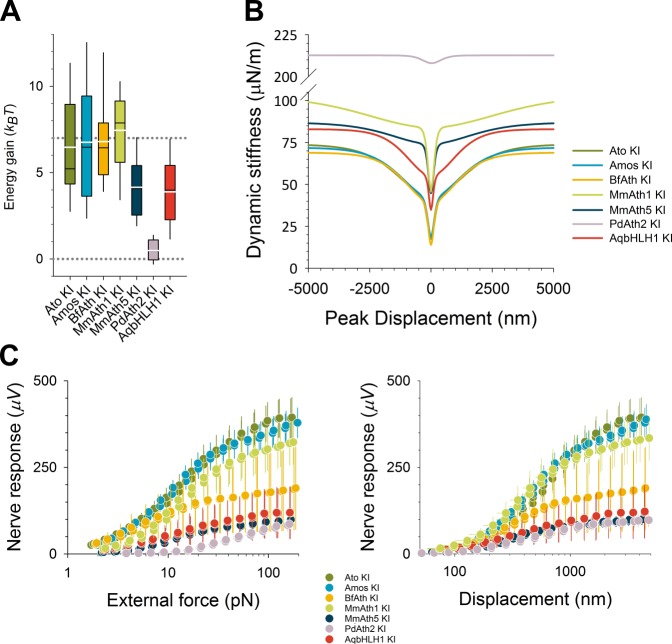
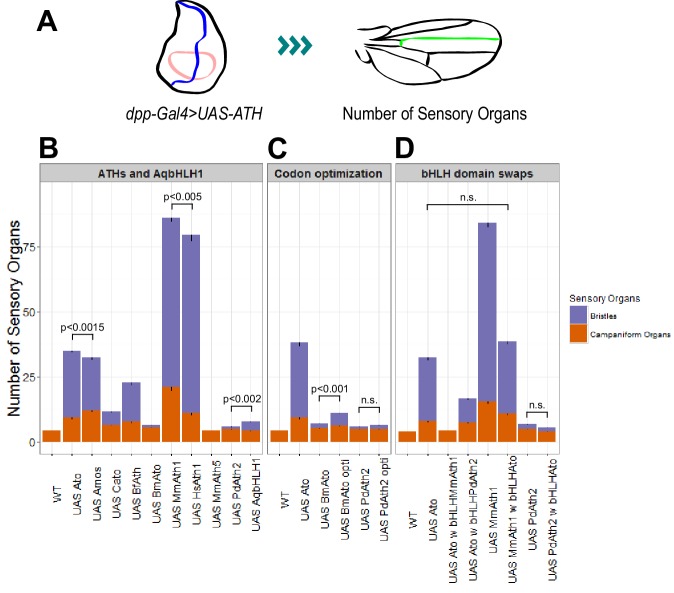
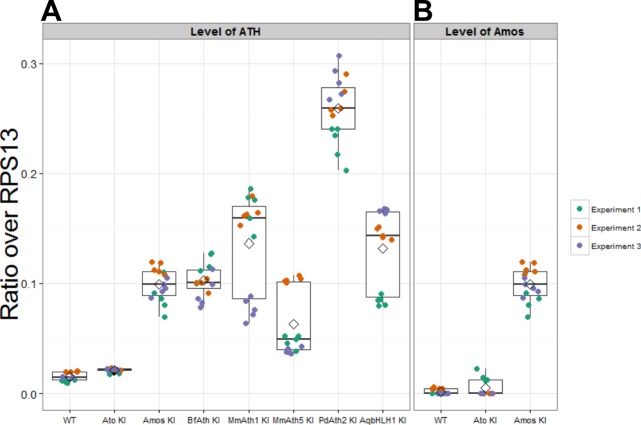
Weinberger Simon, Topping Matthew P, Yan Jiekun, Claeys Annelies, Geest Natalie De, Ozbay Duru, Hassan Talah, He Xiaoli, Albert Joerg T, Hassan Bassem A, Ramaekers Ariane
VIB Center for the Biology of Disease, VIB, Leuven, Belgium.
Center for Human Genetics, University of Leuven School of Medicine, Leuven, Belgium.
Elife. 2017 Apr 13;6:e26402. doi: 10.7554/eLife.26402.
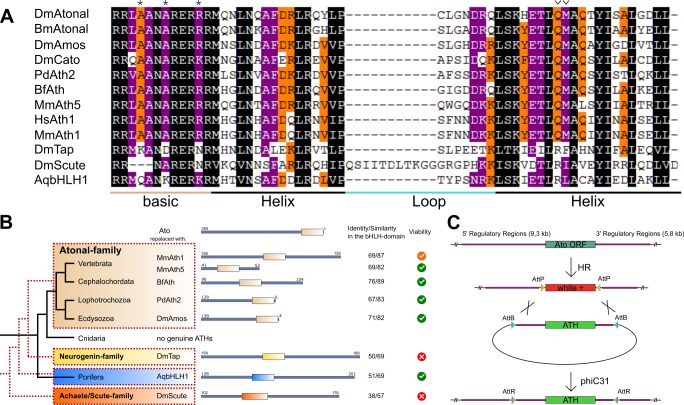
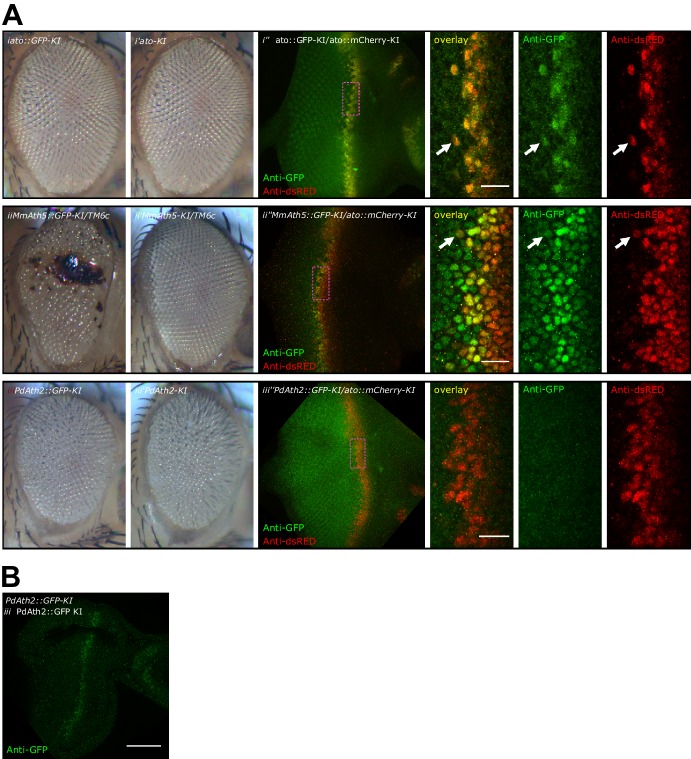
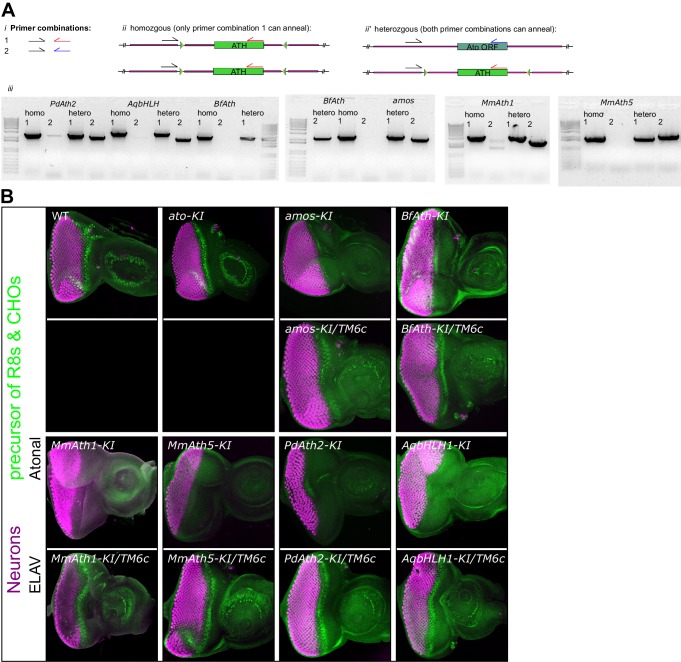
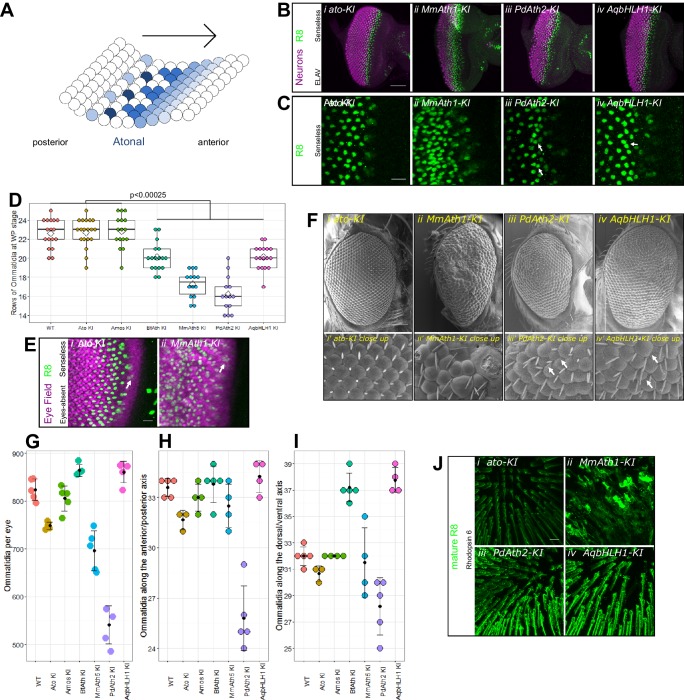
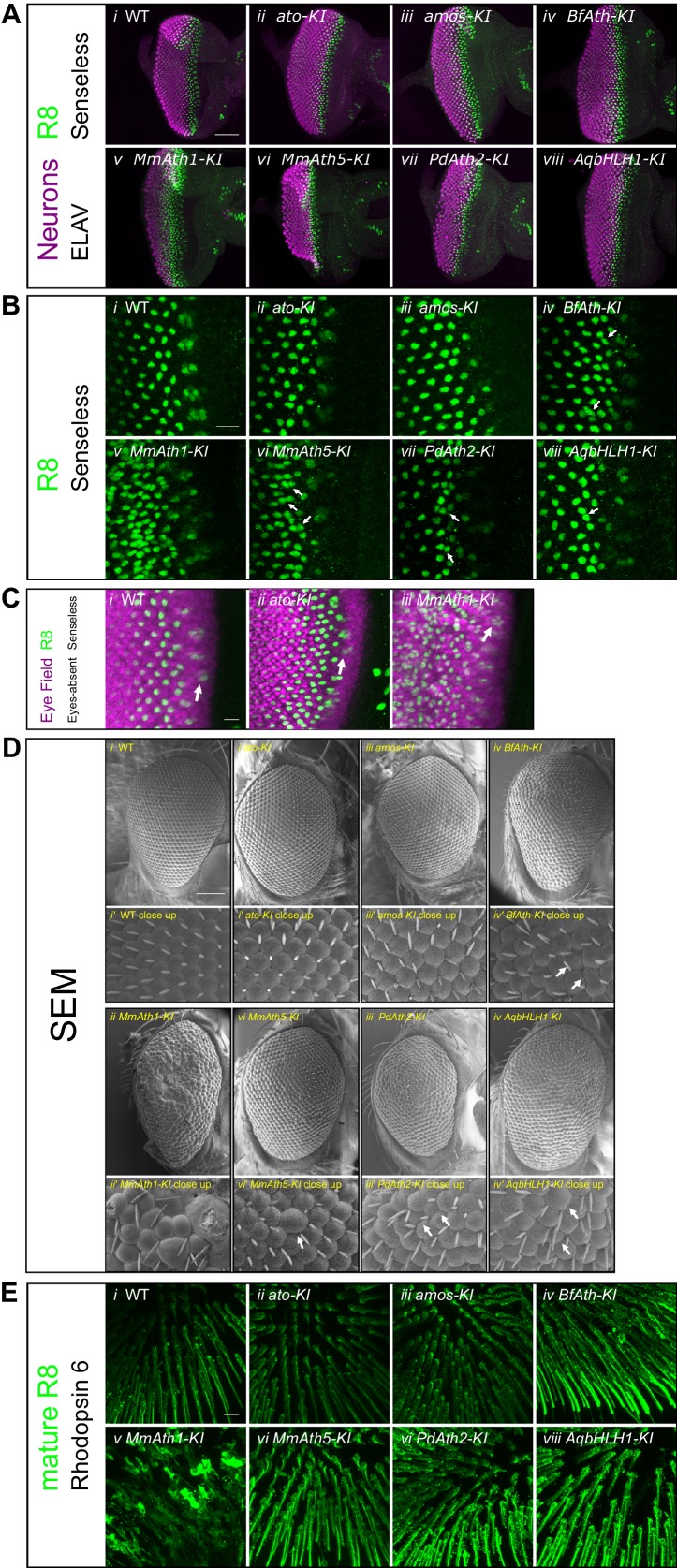
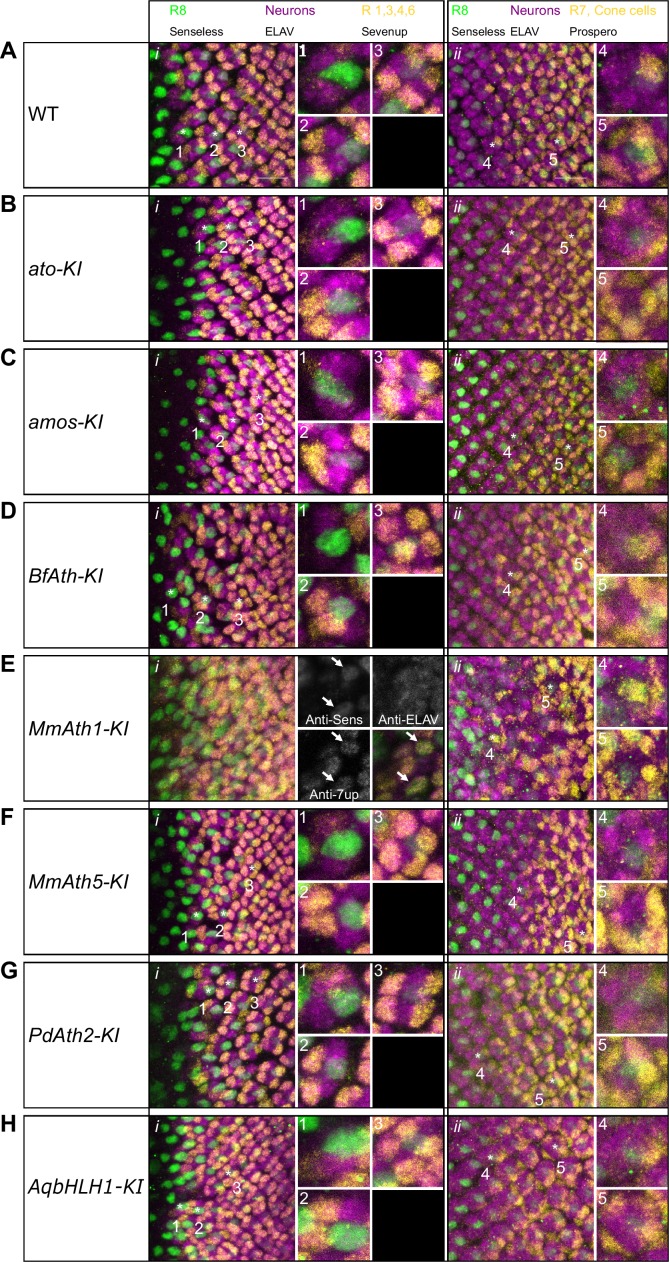
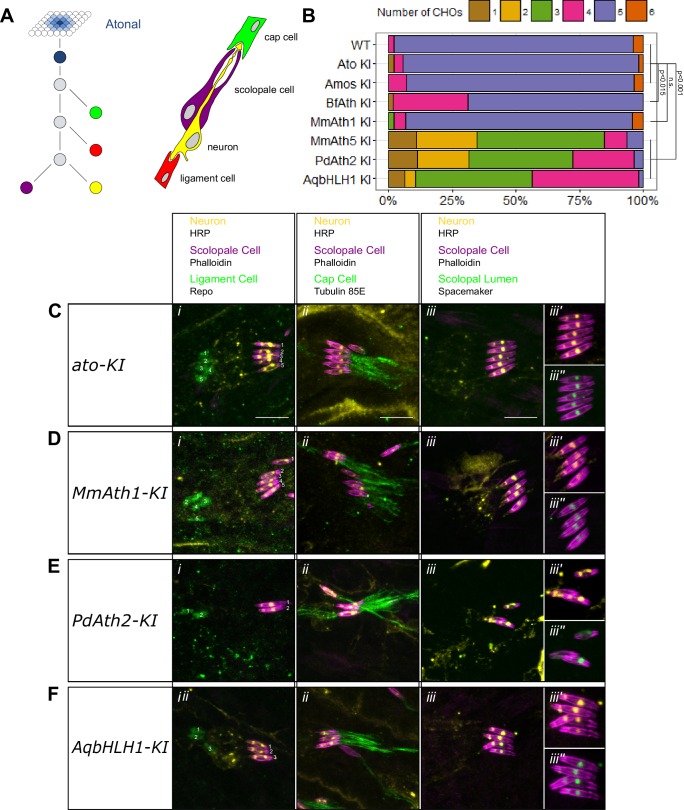
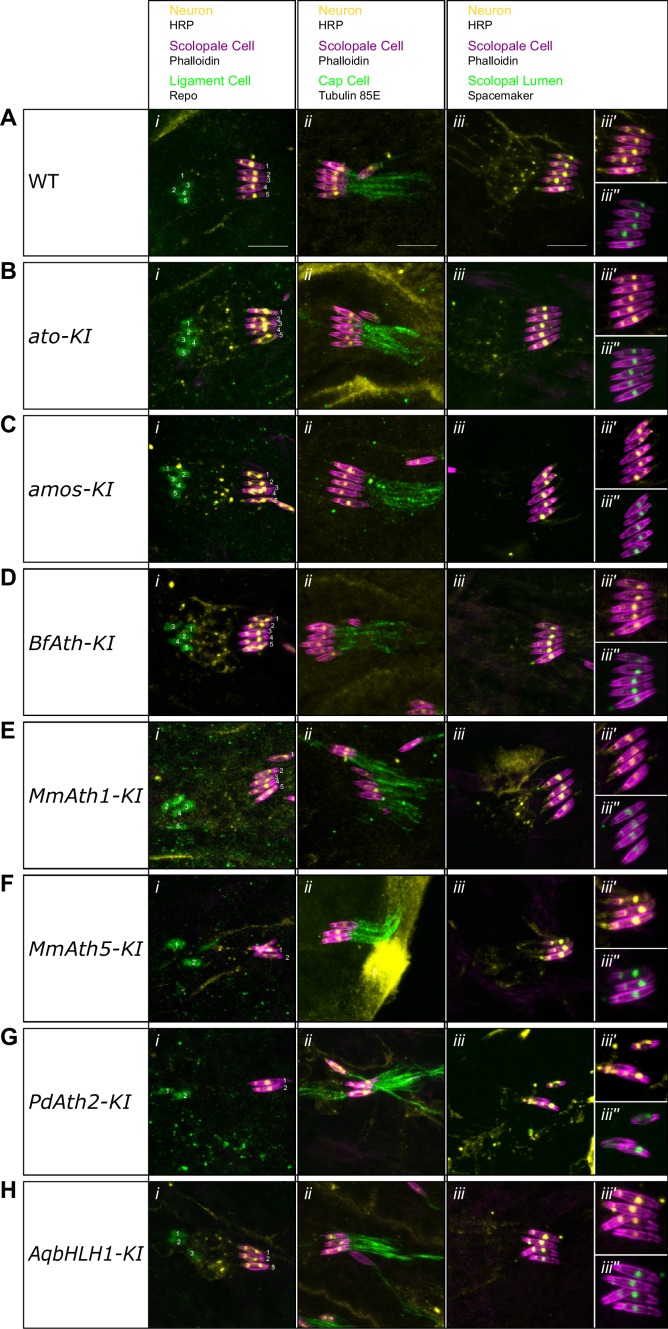
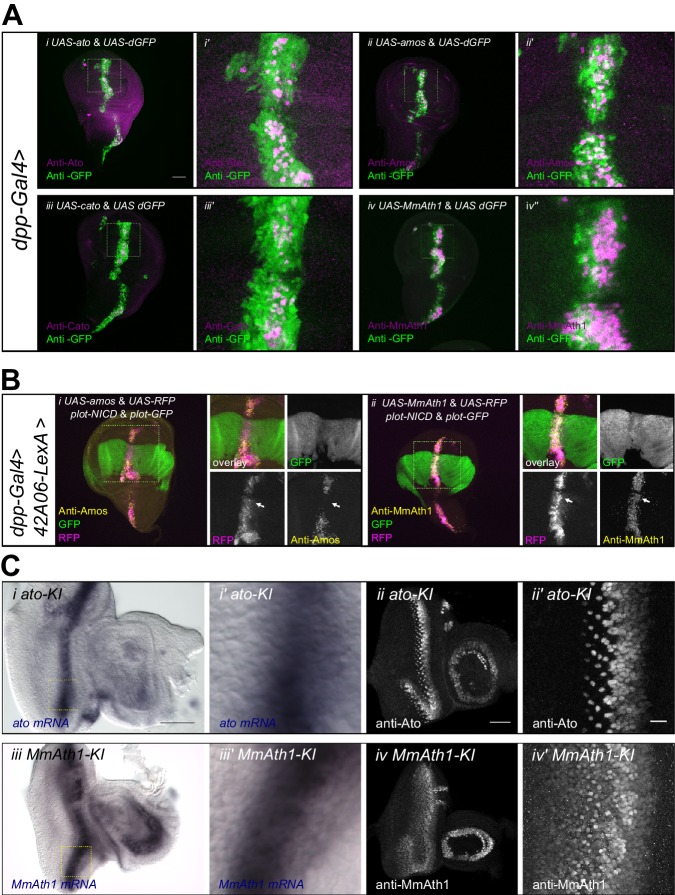
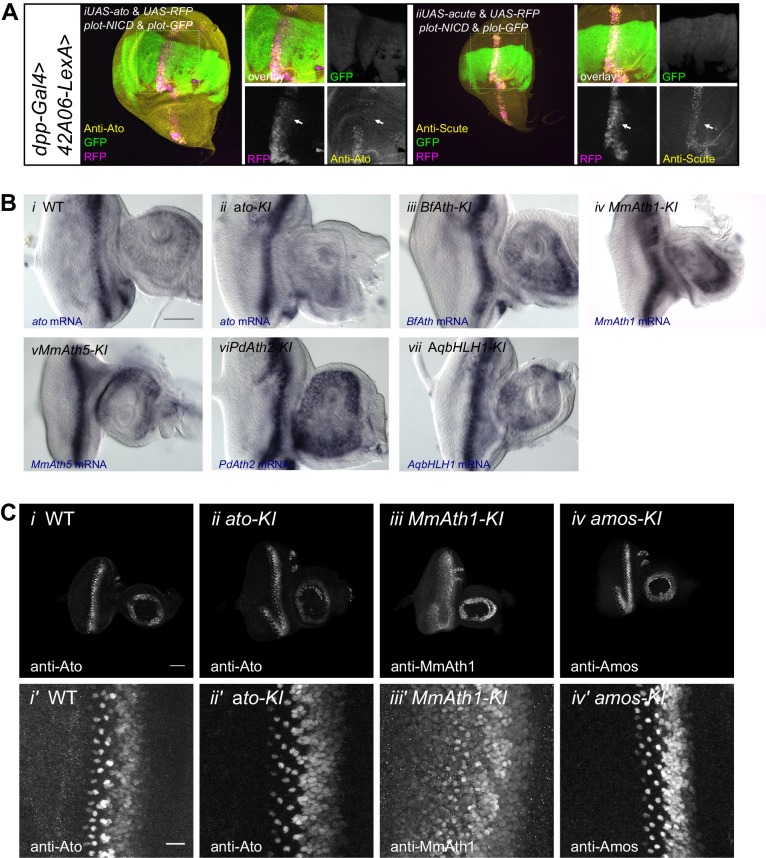
Animals are characterized by a set of highly conserved developmental regulators. Changes in the regulatory elements of these regulators are thought to constitute the major driver of morphological evolution. However, the role of coding sequence evolution remains unresolved. To address this question, we used the Atonal family of proneural transcription factors as a model. coding sequence was endogenously replaced with that of homologues () at key phylogenetic positions, non- proneural genes, and the closest homologue to ancestral proneural genes. and the ancestral-like coding sequences rescued sensory organ fate in mutants, in contrast to non-. Surprisingly, different ATH factors displayed different levels of proneural activity as reflected by the number and functionality of sense organs. This proneural potency gradient correlated directly with ATH protein stability, including in response to Notch signaling, independently of mRNA levels or codon usage. This establishes a distinct and ancient function for ATHs and demonstrates that coding sequence evolution can underlie quantitative variation in sensory development and function.
动物的特征是具有一组高度保守的发育调节因子。这些调节因子的调控元件变化被认为是形态进化的主要驱动力。然而,编码序列进化的作用仍未得到解决。为了解决这个问题,我们以无调性神经前体转录因子家族作为模型。在关键系统发育位置,将编码序列内源性地替换为同源物()、非神经前体基因以及与祖先神经前体基因最接近的同源物的编码序列。与非同源物相比,和类似祖先的编码序列挽救了突变体中的感觉器官命运。令人惊讶的是,不同的无调性神经前体因子显示出不同水平的神经前体活性,这通过感觉器官的数量和功能反映出来。这种神经前体潜能梯度与无调性神经前体蛋白稳定性直接相关,包括对Notch信号的反应,与mRNA水平或密码子使用无关。这确立了无调性神经前体因子独特而古老的功能,并证明编码序列进化可以成为感觉发育和功能定量变异的基础。