Nemoto Yuri L, Morris Roger J, Hijikata Hiroko, Tsunoyama Taka A, Shibata Akihiro C E, Kasai Rinshi S, Kusumi Akihiro, Fujiwara Takahiro K
Center for Meso-Bio Single-Molecule Imaging (CeMI), Institute for Integrated Cell-Material Sciences (WPI-iCeMS), Kyoto University, Kyoto, 606-8507, Japan.
Institute for Frontier Life and Medical Sciences, Kyoto University, Kyoto, 606-8507, Japan.
Cell Biochem Biophys. 2017 Dec;75(3-4):399-412. doi: 10.1007/s12013-017-0808-3. Epub 2017 Jun 24.
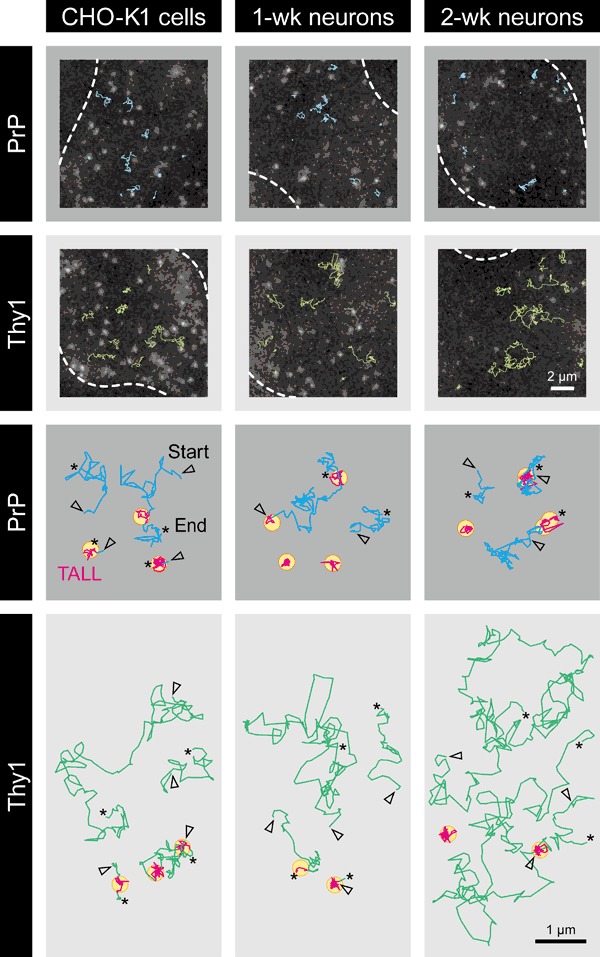
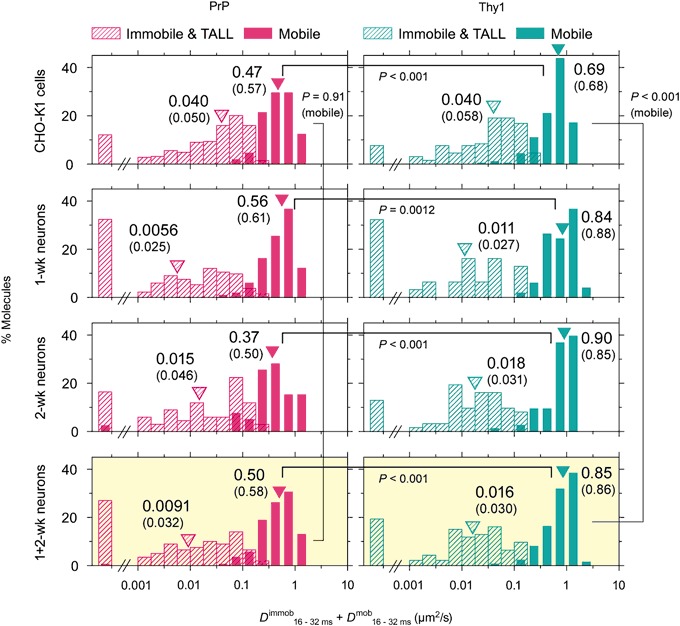
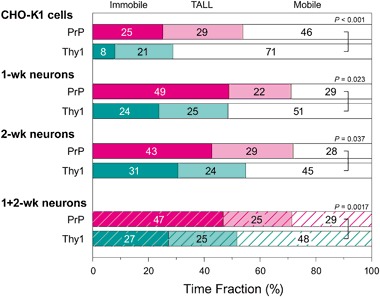
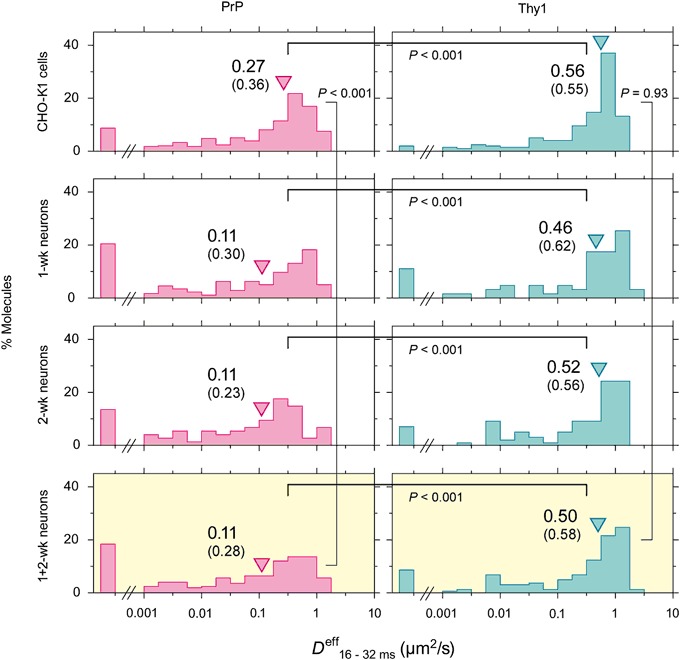
The central mechanism for the transmission of the prion protein misfolding is the structural conversion of the normal cellular prion protein to the pathogenic misfolded prion protein, by the interaction with misfolded prion protein. This process might be enhanced due to the homo-dimerization/oligomerization of normal prion protein. However, the behaviors of normal prion protein in the plasma membrane have remained largely unknown. Here, using single fluorescent-molecule imaging, we found that both prion protein and Thy1, a control glycosylphosphatidylinositol-anchored protein, exhibited very similar intermittent transient immobilizations lasting for a few seconds within an area of 24.2 and 3.5 nm in diameter in CHO-K1 and hippocampal neurons cultured for 1- and 2-weeks, respectively. Prion protein molecules were immobile during 72% of the time, approximately 1.4× more than Thy1, due to prion protein's higher immobilization frequency. When mobile, prion protein diffused 1.7× slower than Thy1. Prion protein's slower diffusion might be caused by its transient interaction with other prion protein molecules, whereas its brief immobilization might be due to temporary association with prion protein clusters. Prion protein molecules might be newly recruited to prion protein clusters all the time, and simultaneously, prion protein molecules in the cluster might be departing continuously. Such dynamic interactions of normal prion protein molecules would strongly enhance the spreading of misfolded prion protein.
朊病毒蛋白错误折叠传播的核心机制是正常细胞朊病毒蛋白通过与错误折叠的朊病毒蛋白相互作用,发生结构转变成为致病性错误折叠的朊病毒蛋白。由于正常朊病毒蛋白的同二聚化/寡聚化,这一过程可能会增强。然而,正常朊病毒蛋白在质膜中的行为在很大程度上仍不清楚。在这里,我们使用单荧光分子成像发现,在分别培养1周和2周的CHO-K1细胞和海马神经元中,朊病毒蛋白和作为对照的糖基磷脂酰肌醇锚定蛋白Thy1,在直径分别为24.2纳米和3.5纳米的区域内都表现出非常相似的间歇性短暂固定,持续数秒。由于朊病毒蛋白的固定频率更高,朊病毒蛋白分子在72%的时间内是固定的,大约是Thy1的1.4倍。当处于移动状态时,朊病毒蛋白的扩散速度比Thy1慢1.7倍。朊病毒蛋白较慢的扩散可能是由于其与其他朊病毒蛋白分子的短暂相互作用,而其短暂的固定可能是由于与朊病毒蛋白簇的临时结合。朊病毒蛋白分子可能一直在新招募到朊病毒蛋白簇中,同时,簇中的朊病毒蛋白分子可能在不断离开。正常朊病毒蛋白分子的这种动态相互作用将强烈增强错误折叠的朊病毒蛋白的传播。