Epigenetic Discovery, China Novartis Institutes for BioMedical Research, Shanghai, China.
Department of Bioinformatics, Tongji University, Shanghai, China.
Elife. 2017 Aug 2;6:e26129. doi: 10.7554/eLife.26129.
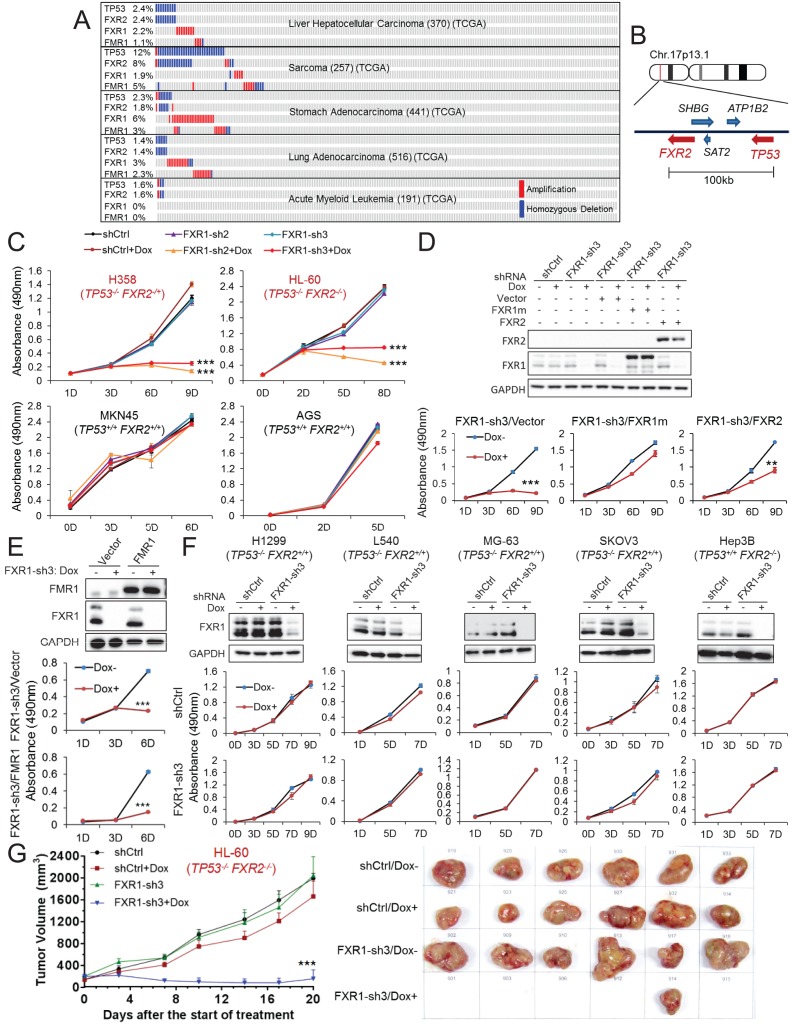
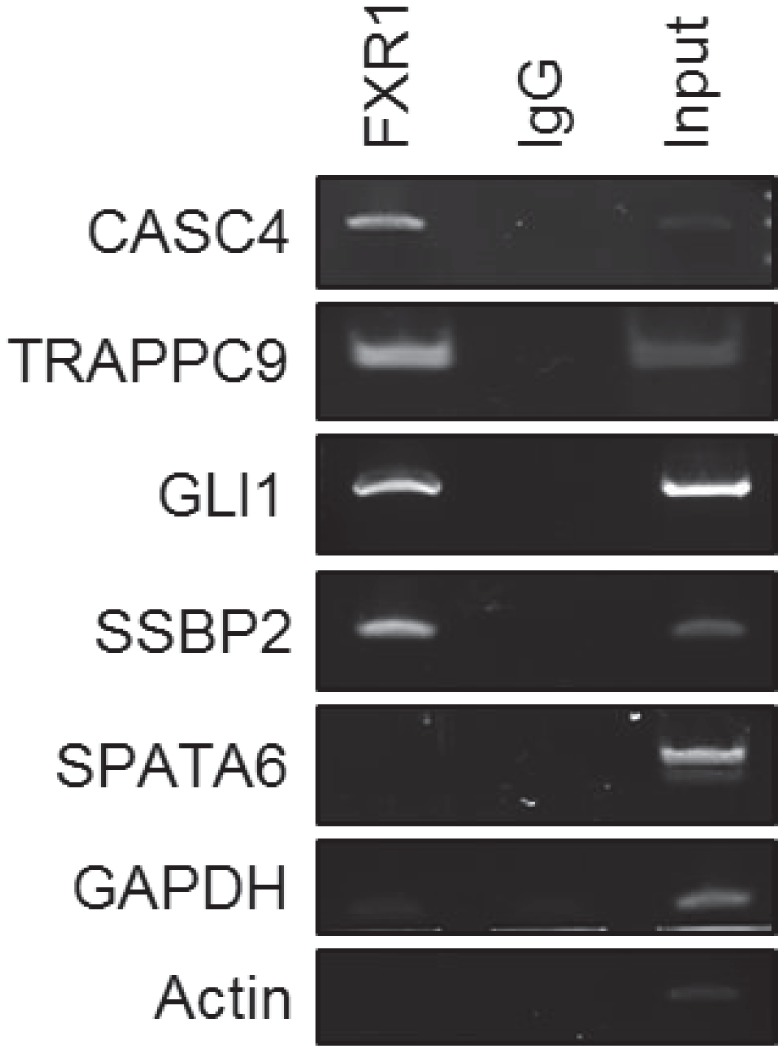
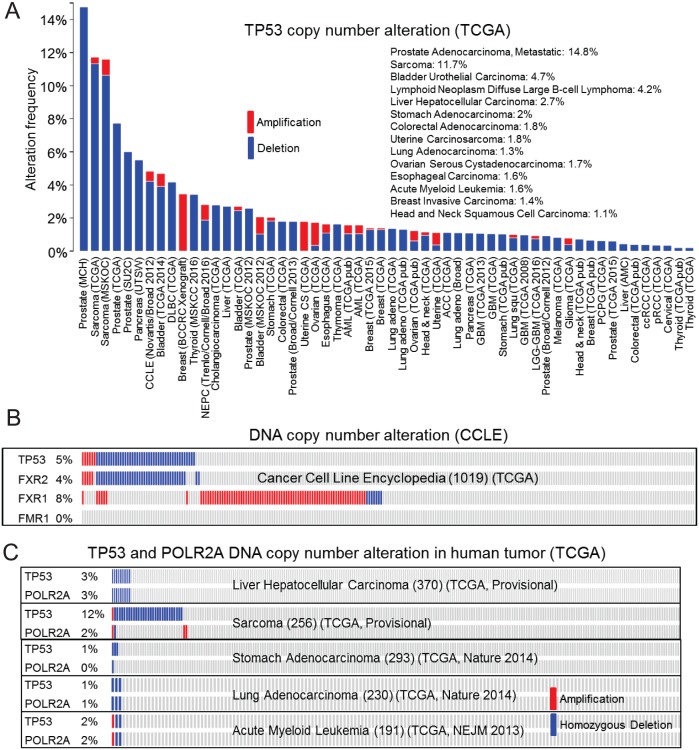
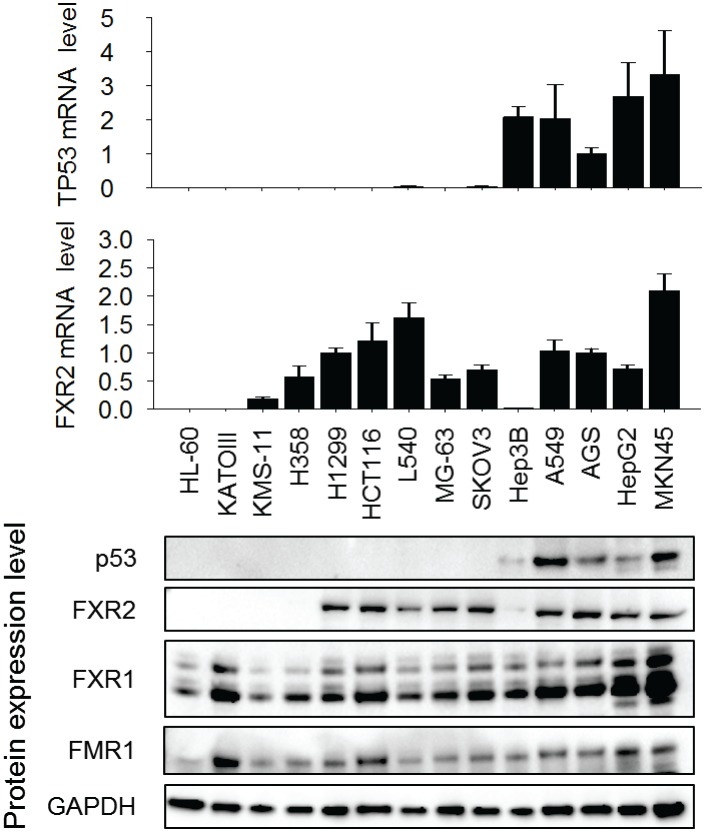
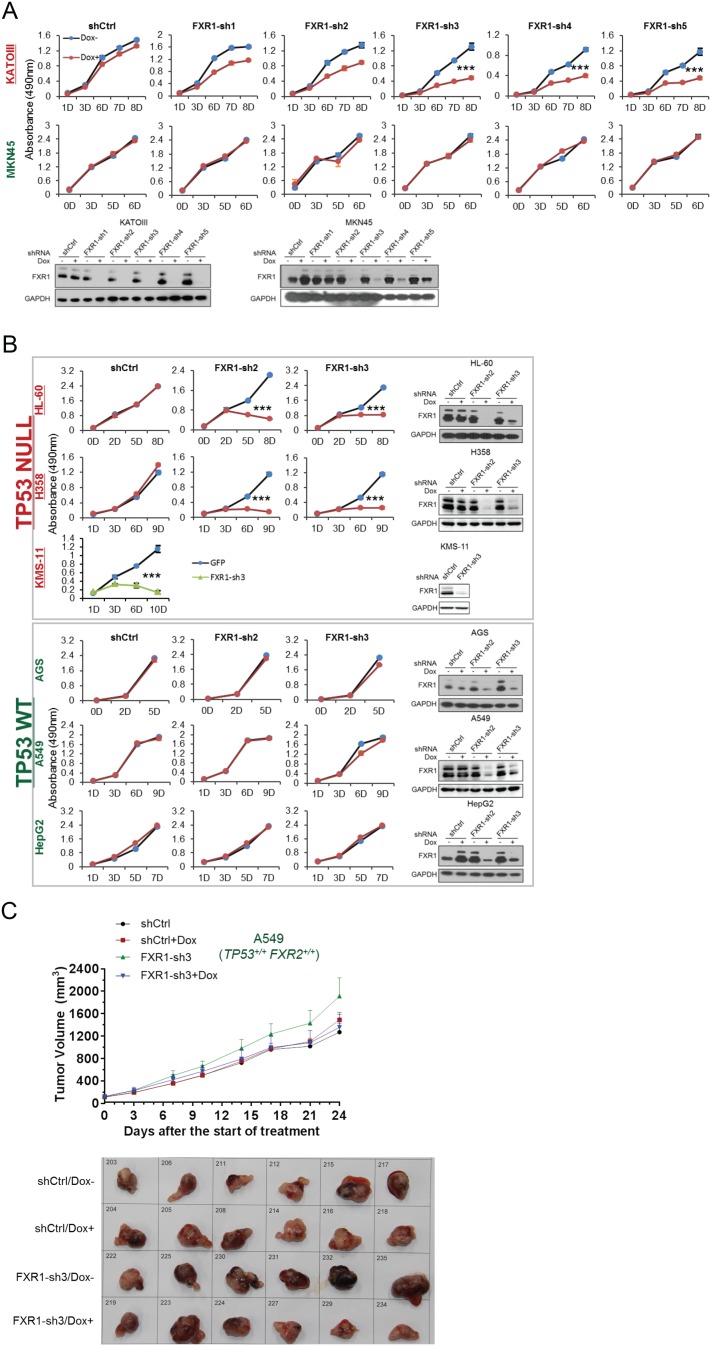
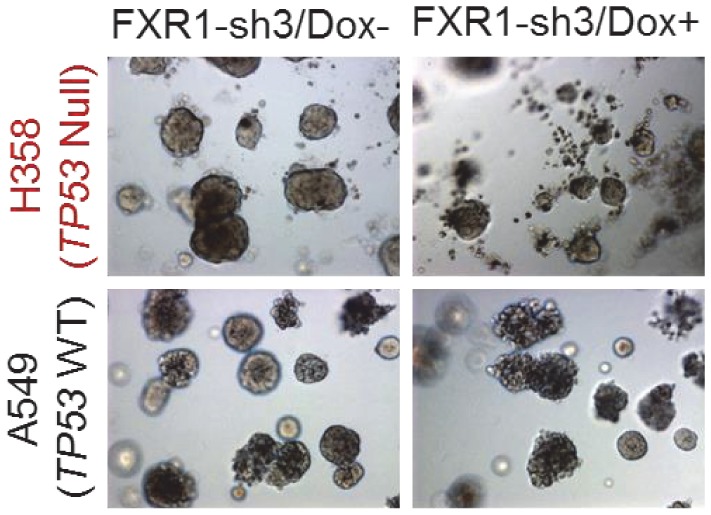
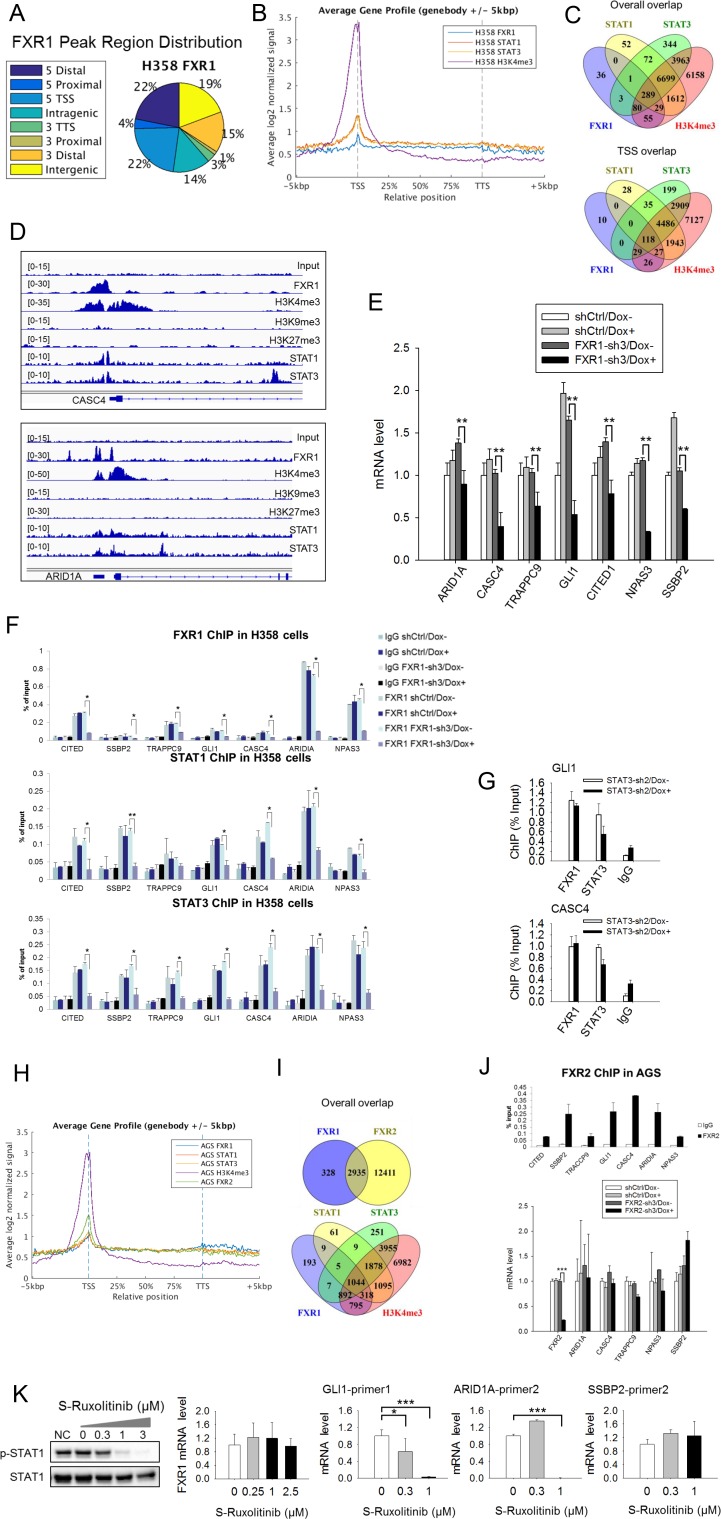
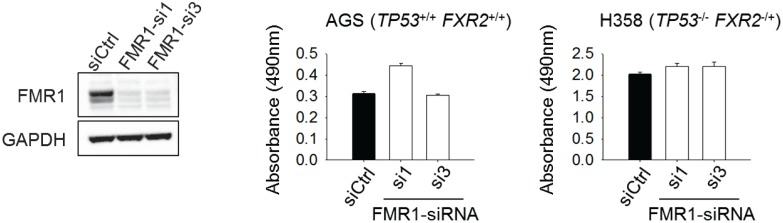
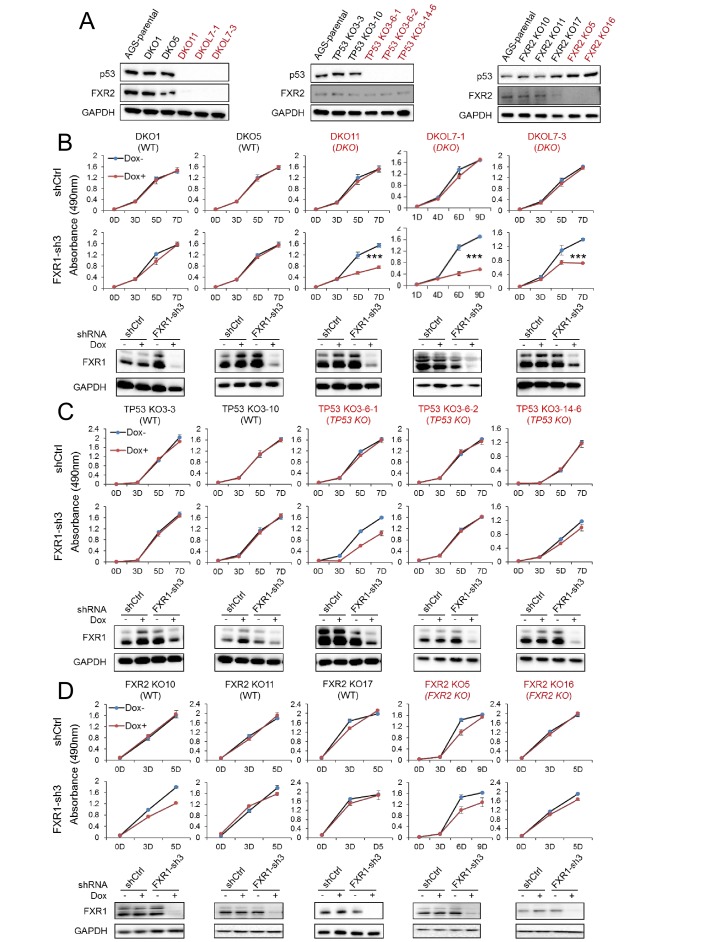
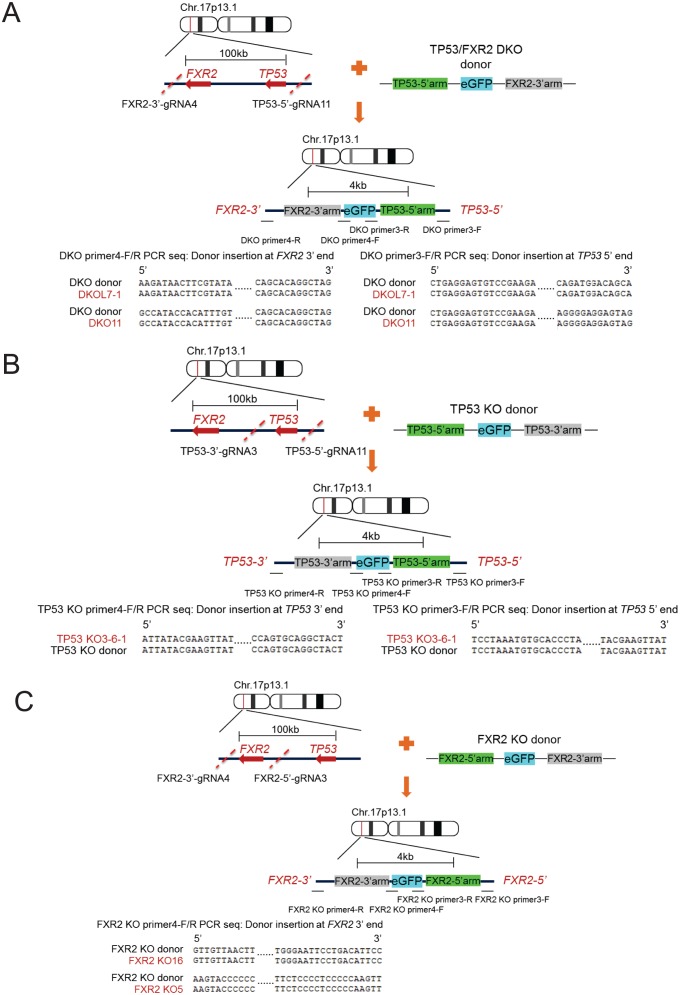
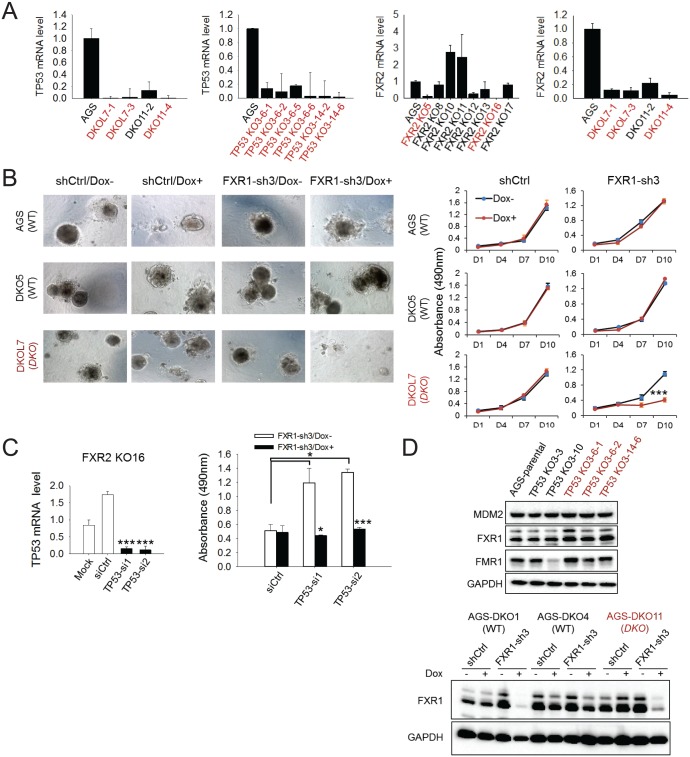
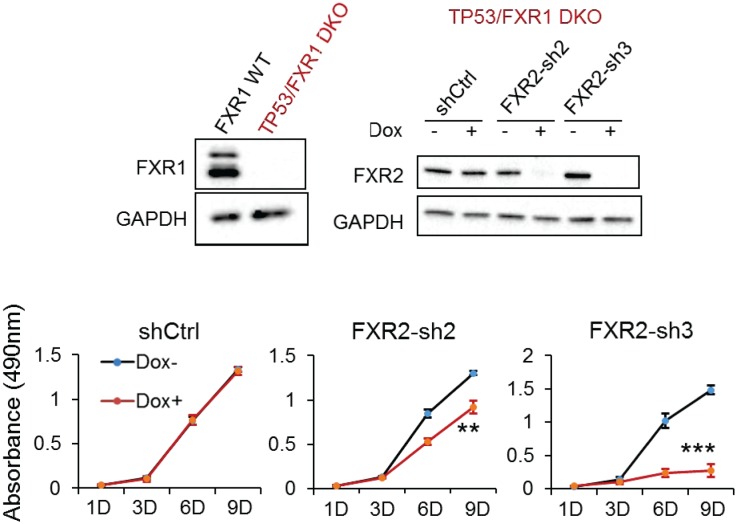
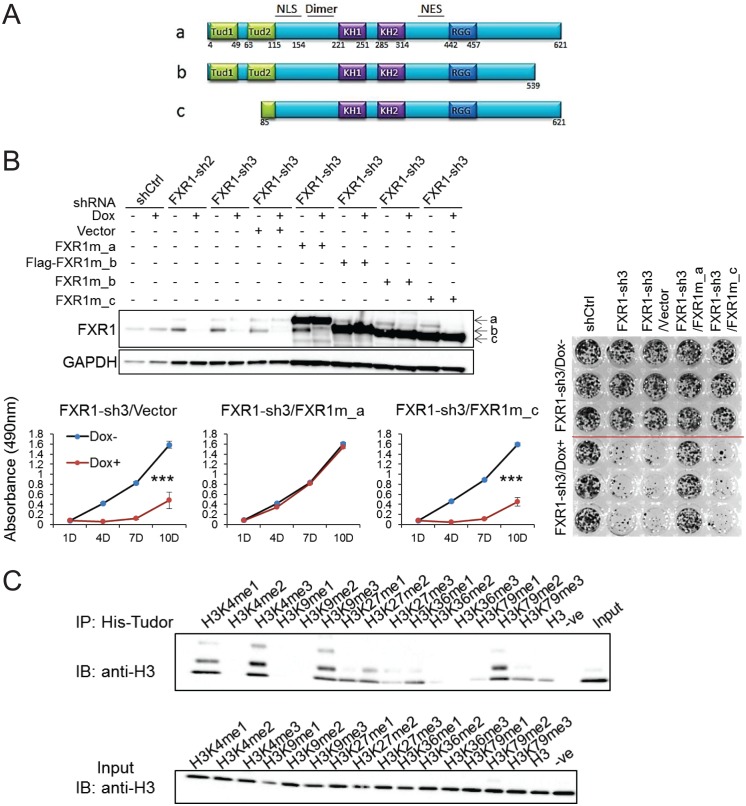
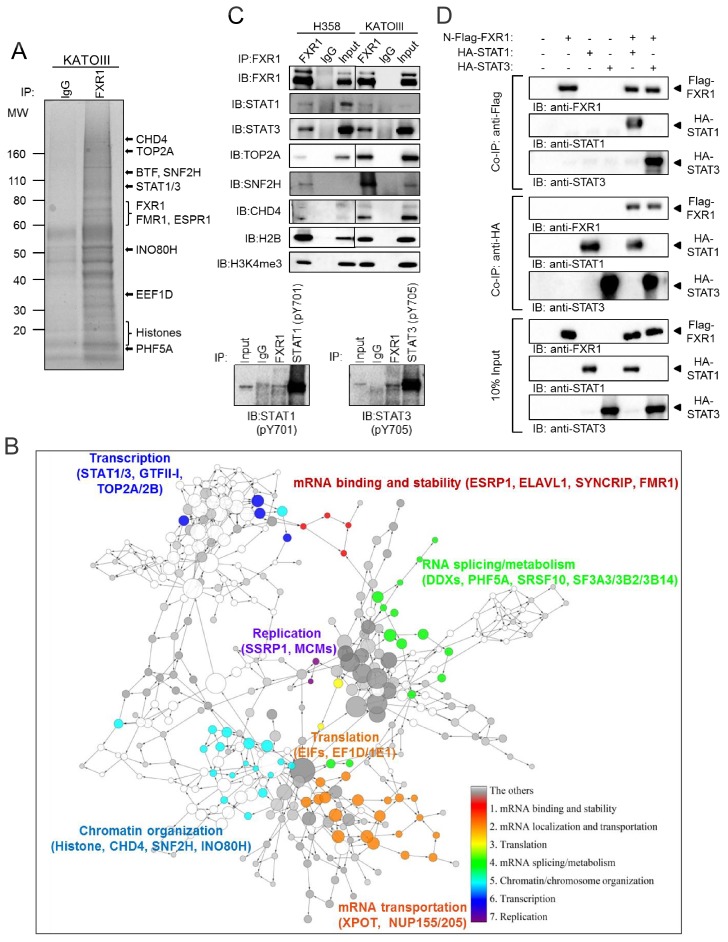
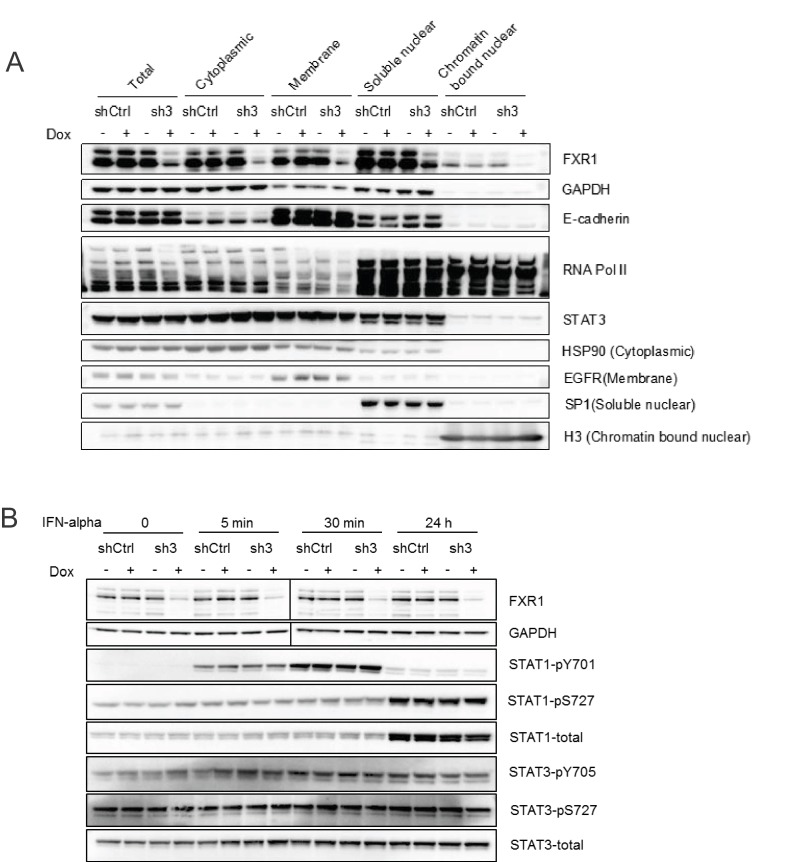
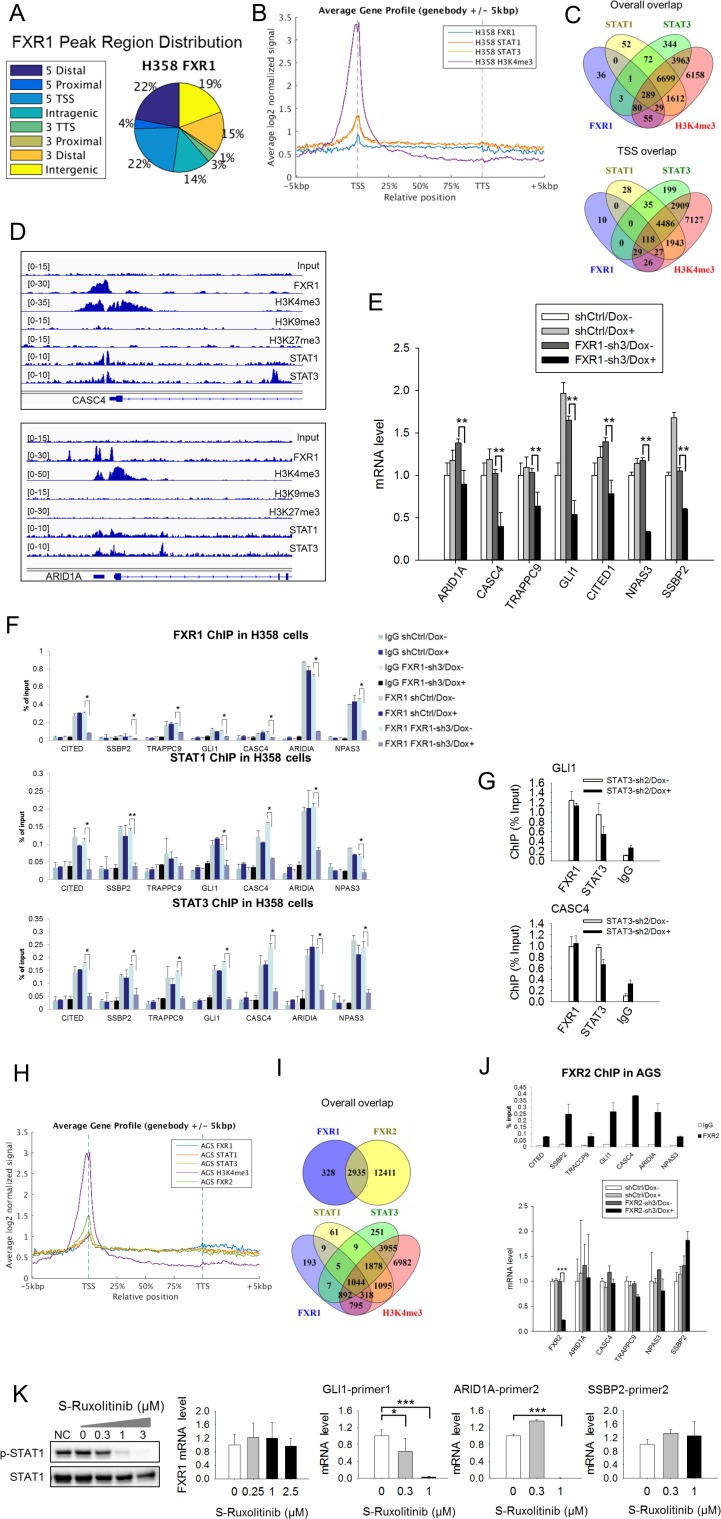
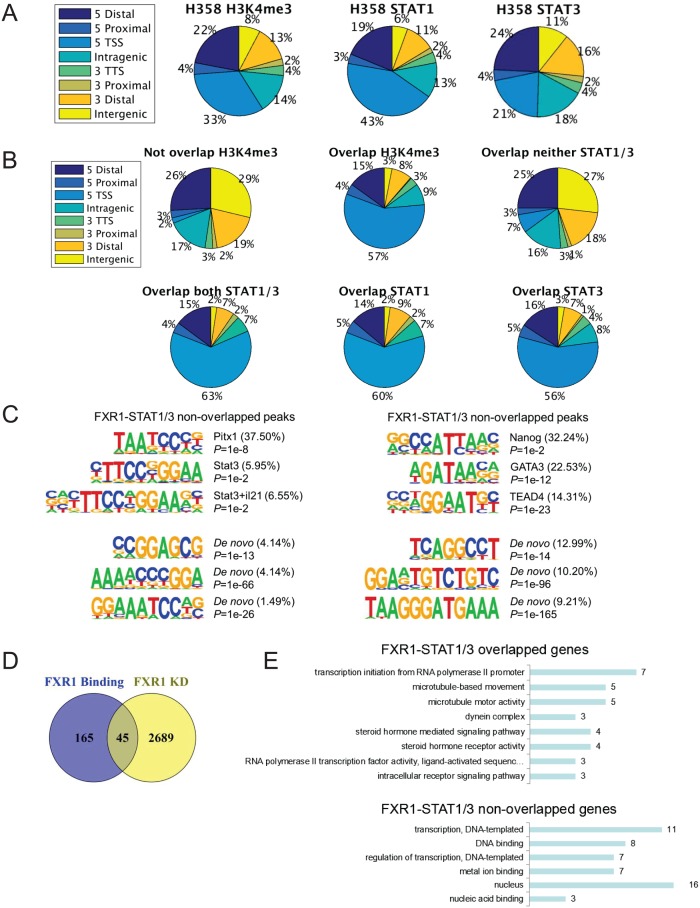
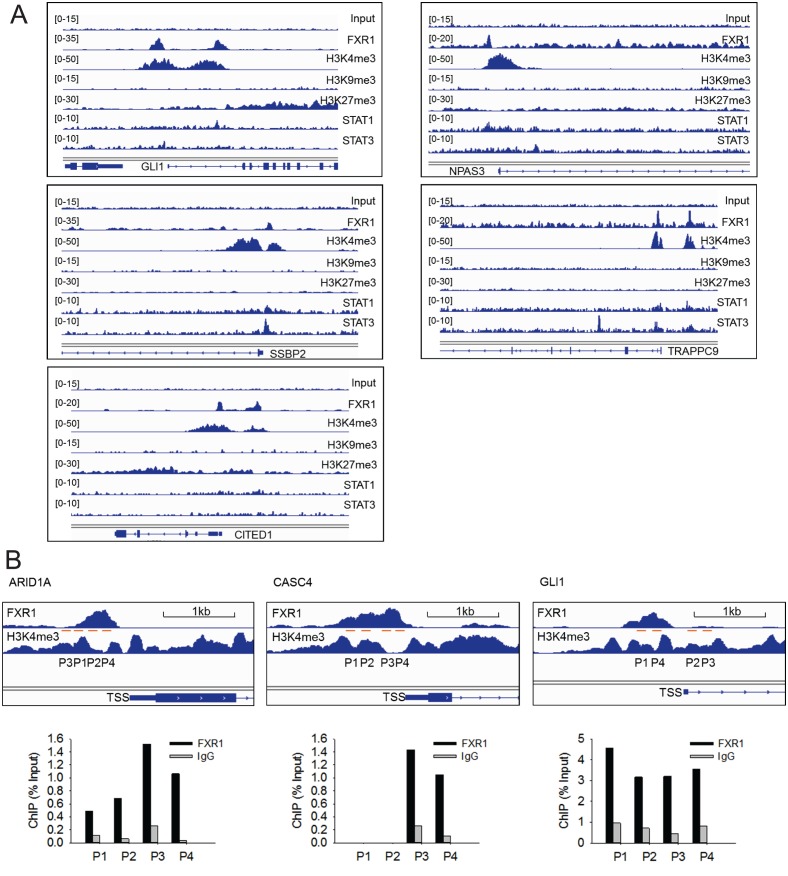
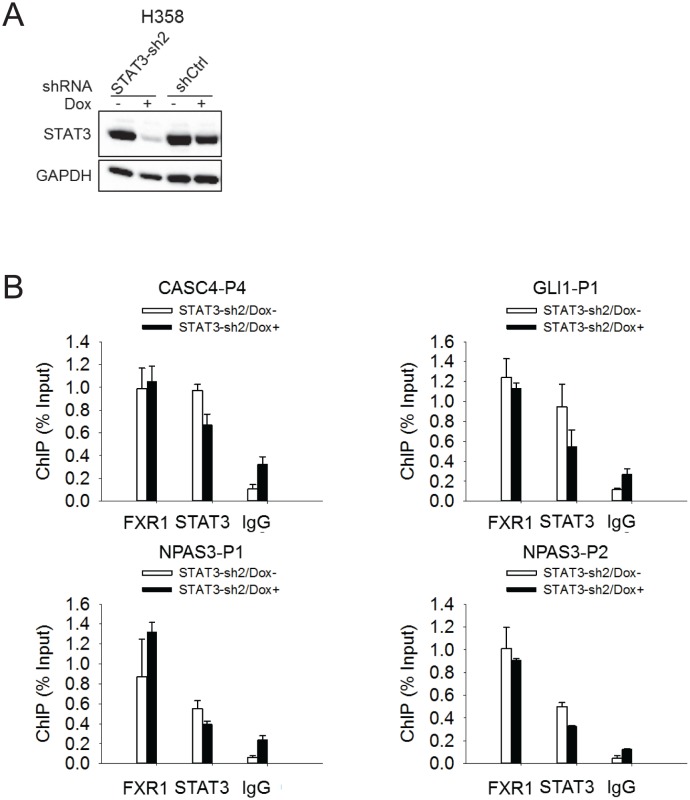
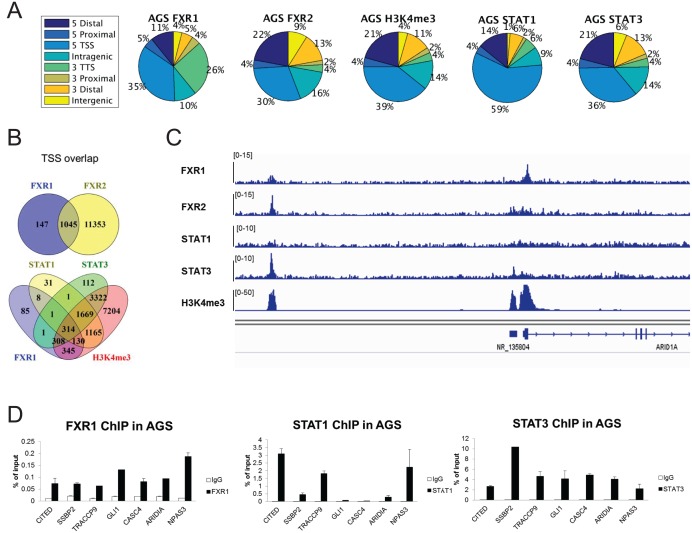
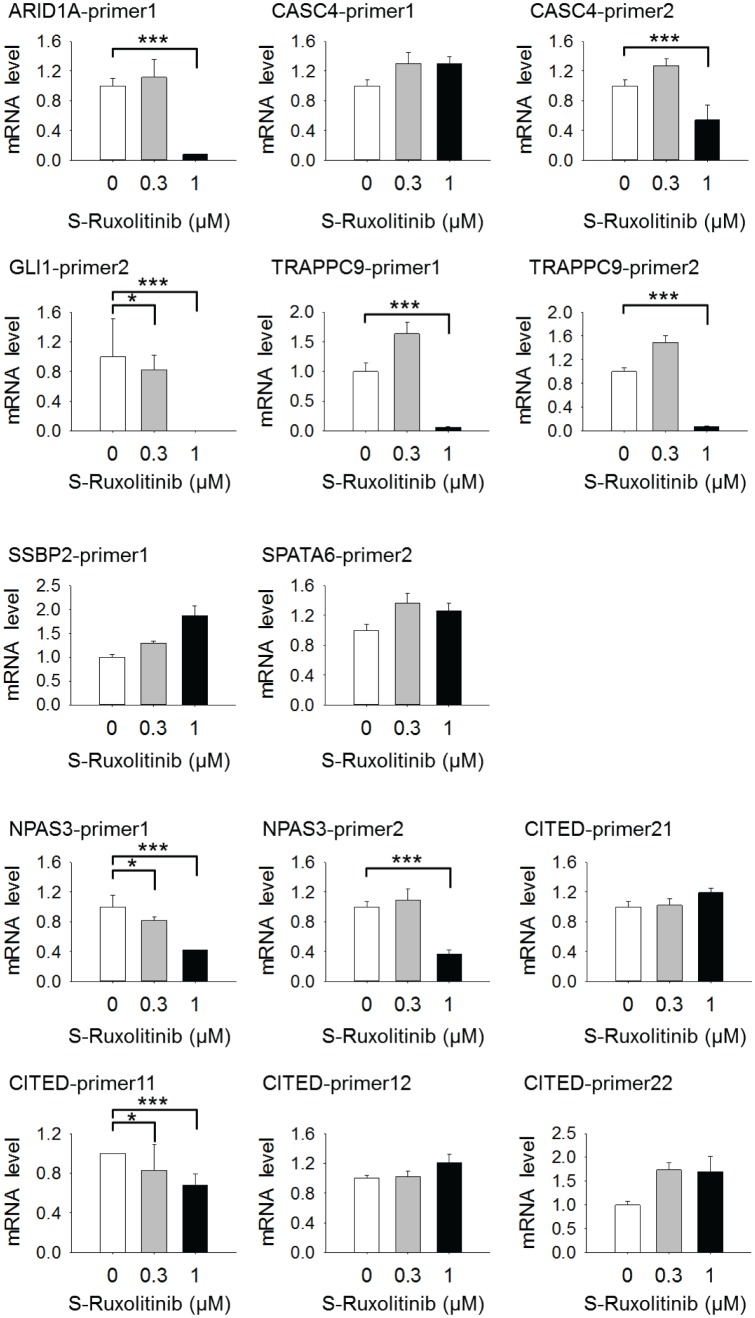
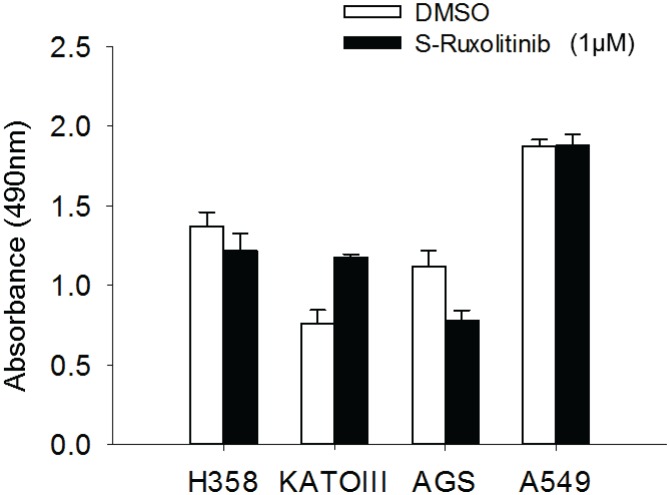
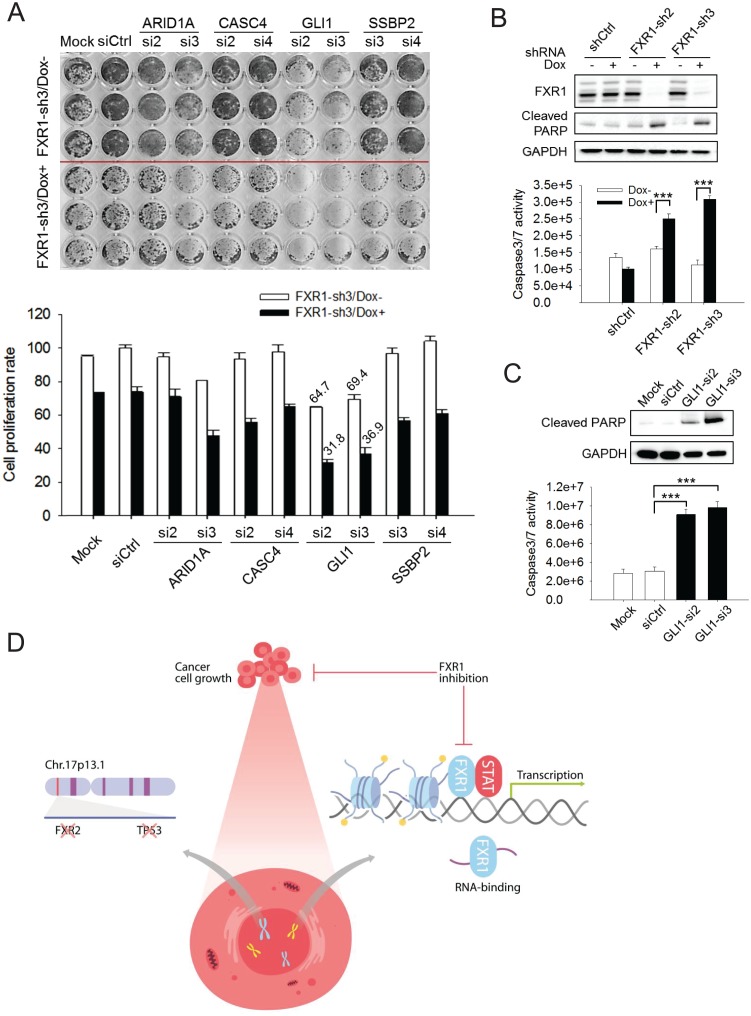
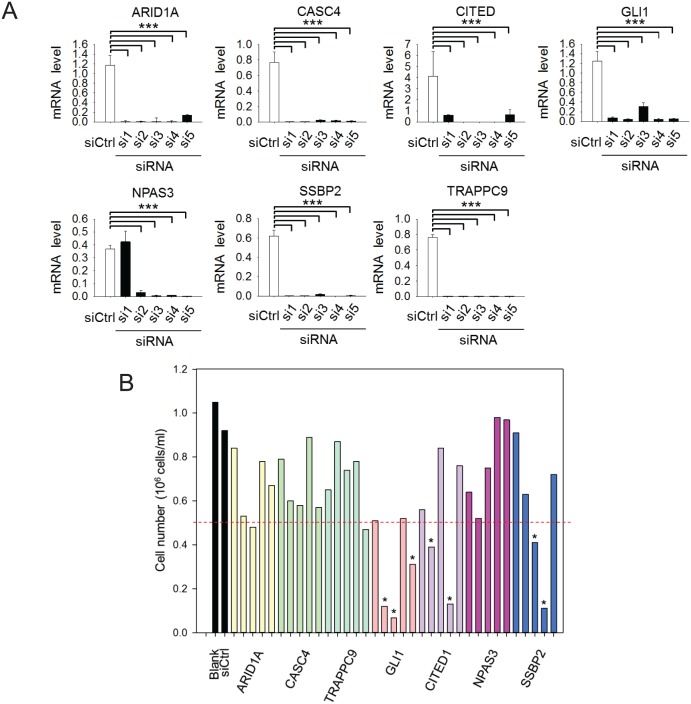
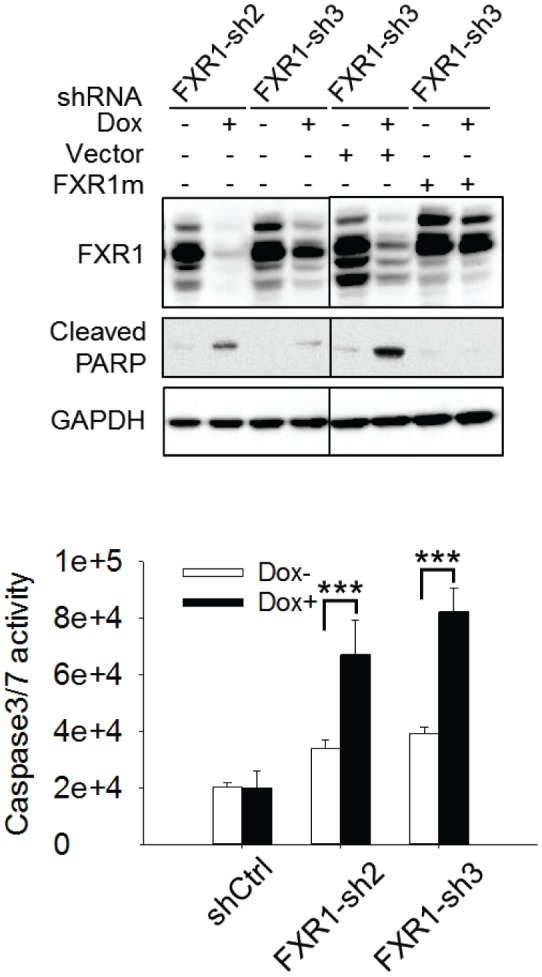
Tumor suppressor p53 prevents cell transformation by inducing apoptosis and other responses. Homozygous deletion occurs in various types of human cancers for which no therapeutic strategies have yet been reported. TCGA database analysis shows that the homozygous deletion locus mostly exhibits co-deletion of the neighboring gene which belongs to the Fragile X gene family. Here, we demonstrate that inhibition of the remaining family member FXR1 selectively blocks cell proliferation in human cancer cells containing homozygous deletion of both and in a collateral lethality manner. Mechanistically, in addition to its RNA-binding function, FXR1 recruits transcription factor STAT1 or STAT3 to gene promoters at the chromatin interface and regulates transcription thus, at least partially, mediating cell proliferation. Our study anticipates that inhibition of FXR1 is a potential therapeutic approach to targeting human cancers harboring homozygous deletion.
抑癌基因 p53 通过诱导细胞凋亡和其他反应来阻止细胞转化。在各种类型的人类癌症中都发生了纯合缺失,但目前尚无报道针对这种缺失的治疗策略。TCGA 数据库分析表明,纯合缺失位点大多表现为相邻基因的共缺失,这些基因属于脆性 X 基因家族。在这里,我们证明了抑制剩余的家族成员 FXR1 以旁系致死的方式选择性地阻断含有 和 同时纯合缺失的人类癌细胞的增殖。从机制上讲,除了其 RNA 结合功能外,FXR1 还将转录因子 STAT1 或 STAT3 募集到染色质界面上的基因启动子处,并调节转录,从而至少部分介导细胞增殖。我们的研究预计,抑制 FXR1 可能是针对携带 纯合缺失的人类癌症的潜在治疗方法。