Gorin Aleksandr M, Du Yushen, Liu Franklin Y, Zhang Tian-Hao, Ng Hwee L, Hofmann Christian, Cumberland William G, Sun Ren, Yang Otto O
Department of Microbiology, Immunology & Molecular Genetics, David Geffen School of Medicine, University of California Los Angeles, Los Angeles, California, United States of America.
Department of Molecular and Medical Pharmacology, David Geffen School of Medicine, University of California Los Angeles, Los Angeles, California, United States of America.
PLoS Pathog. 2017 Aug 7;13(8):e1006541. doi: 10.1371/journal.ppat.1006541. eCollection 2017 Aug.
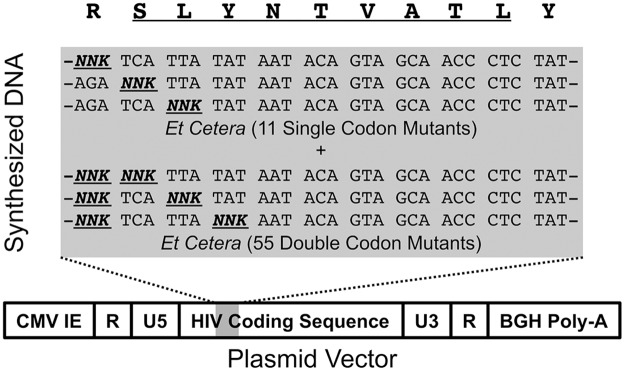
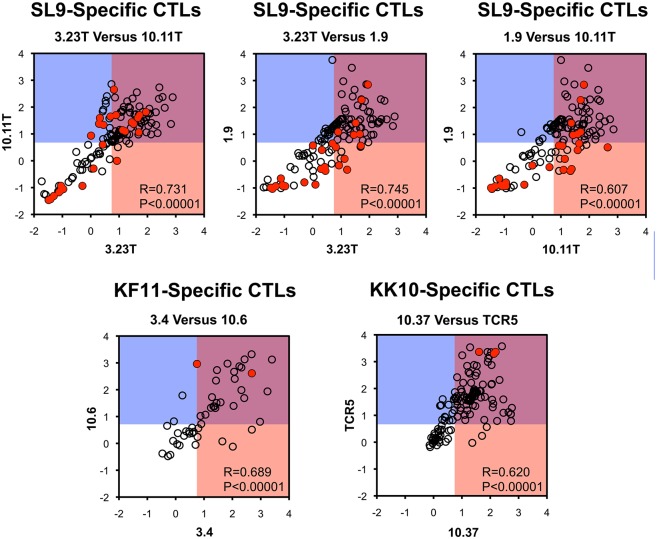
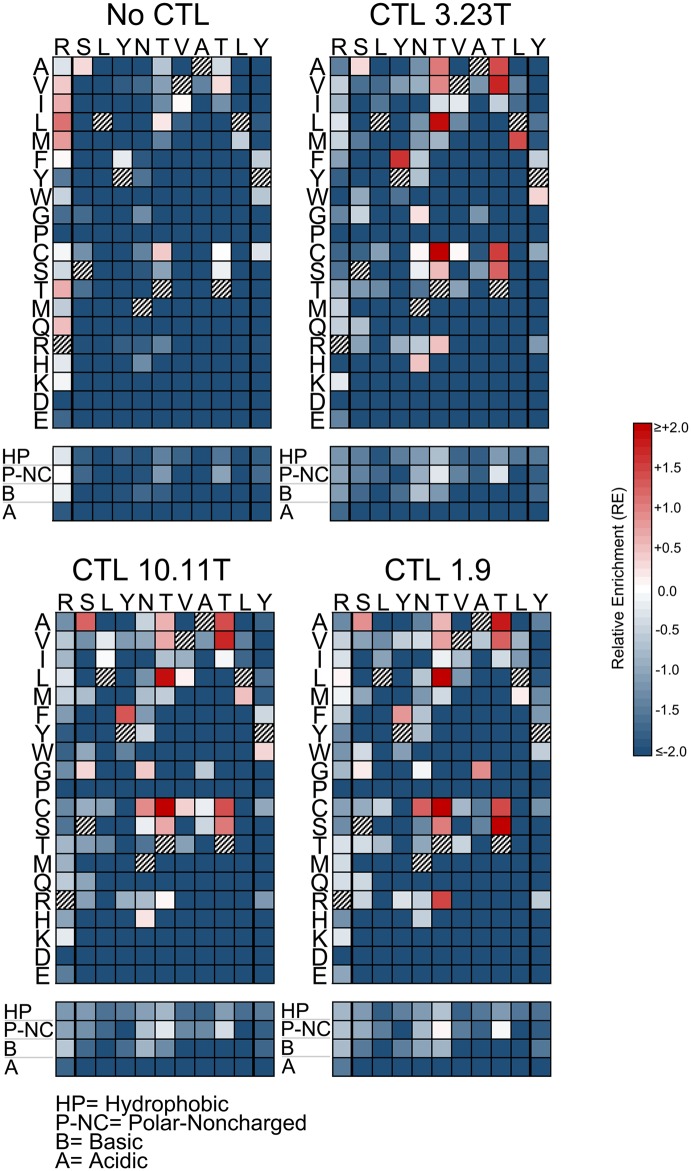
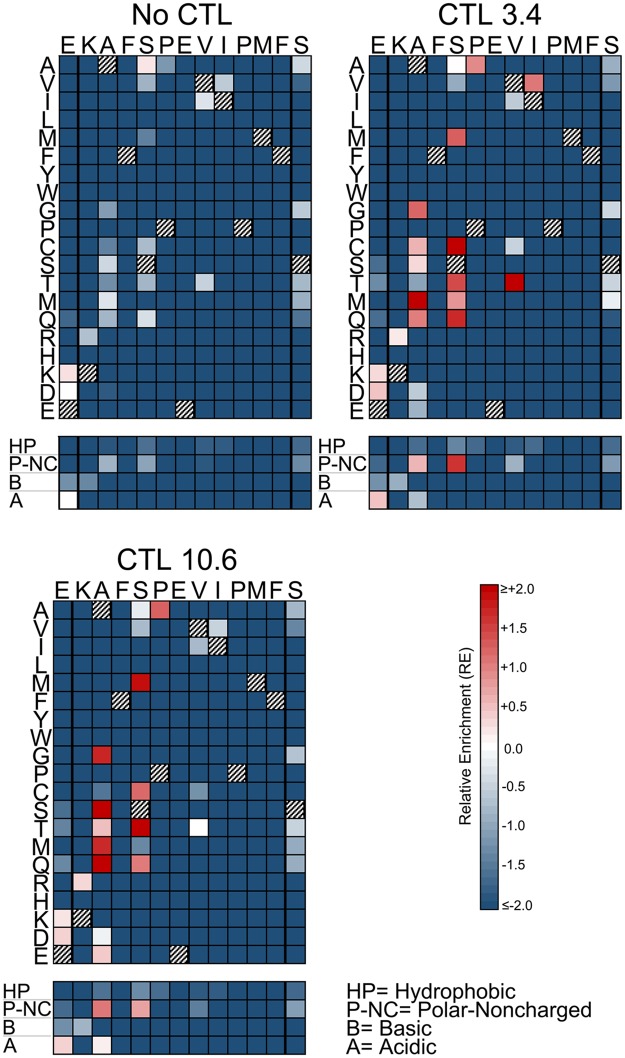
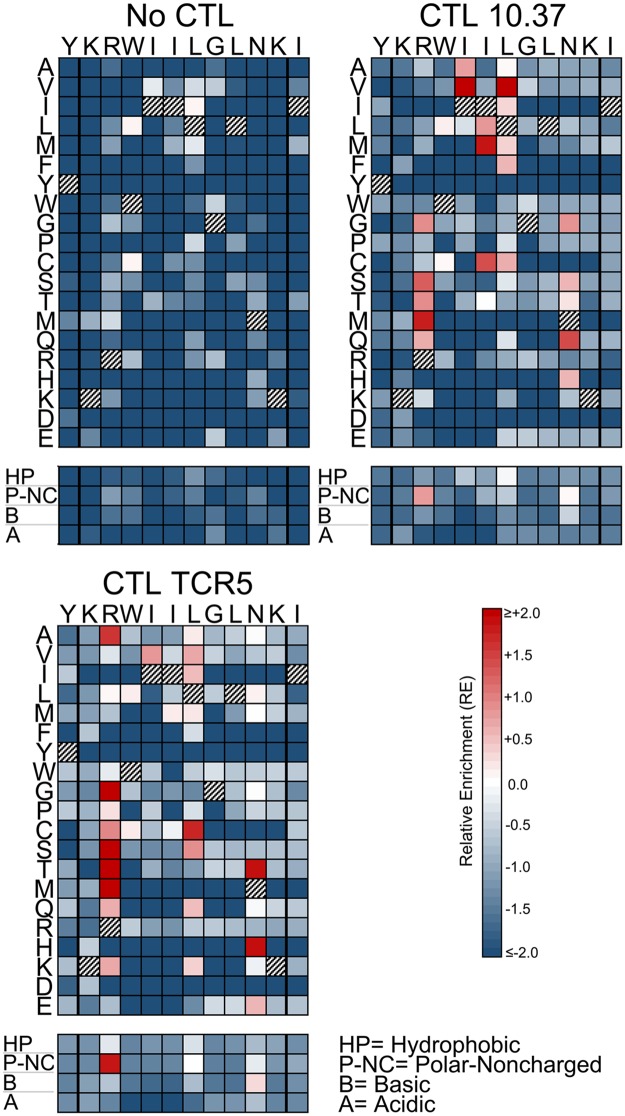
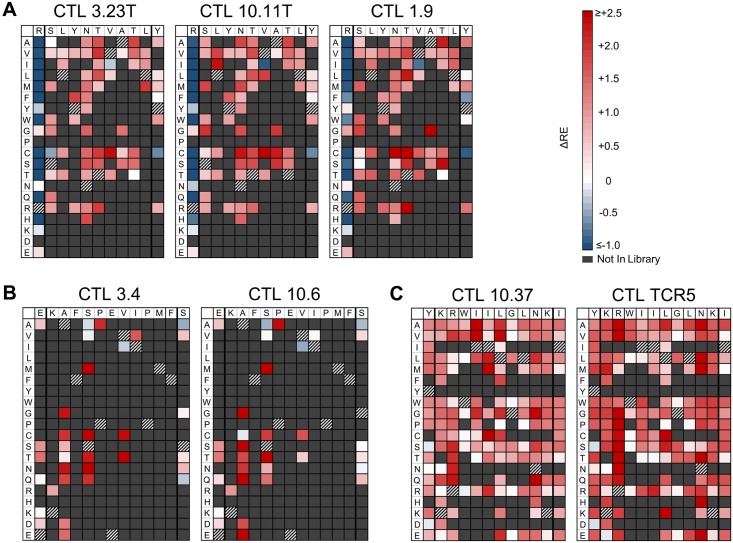
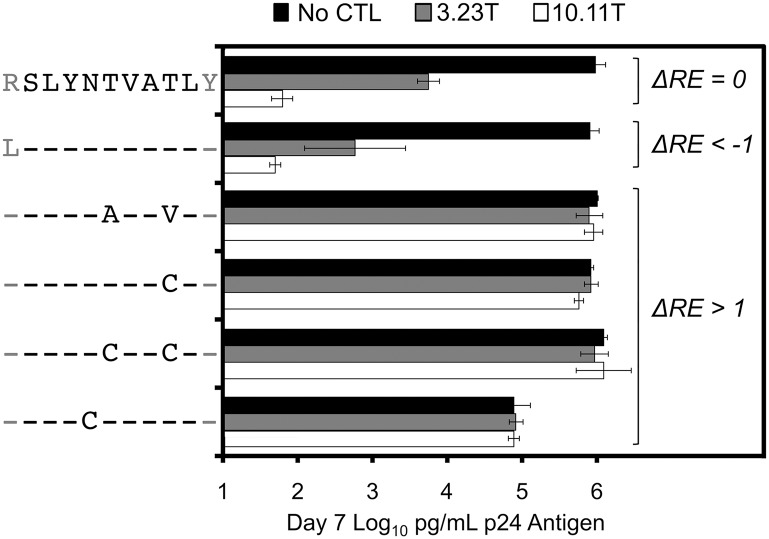
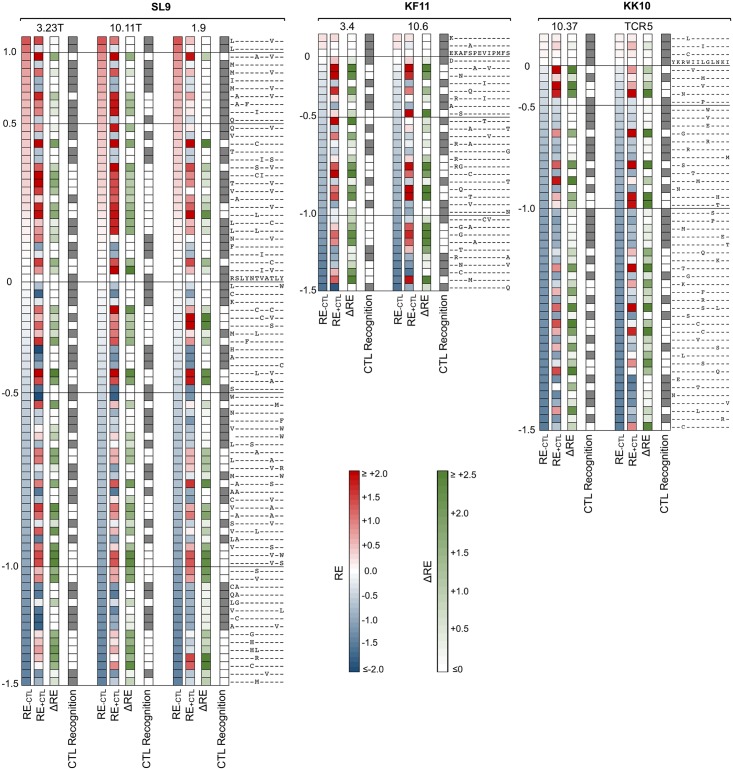
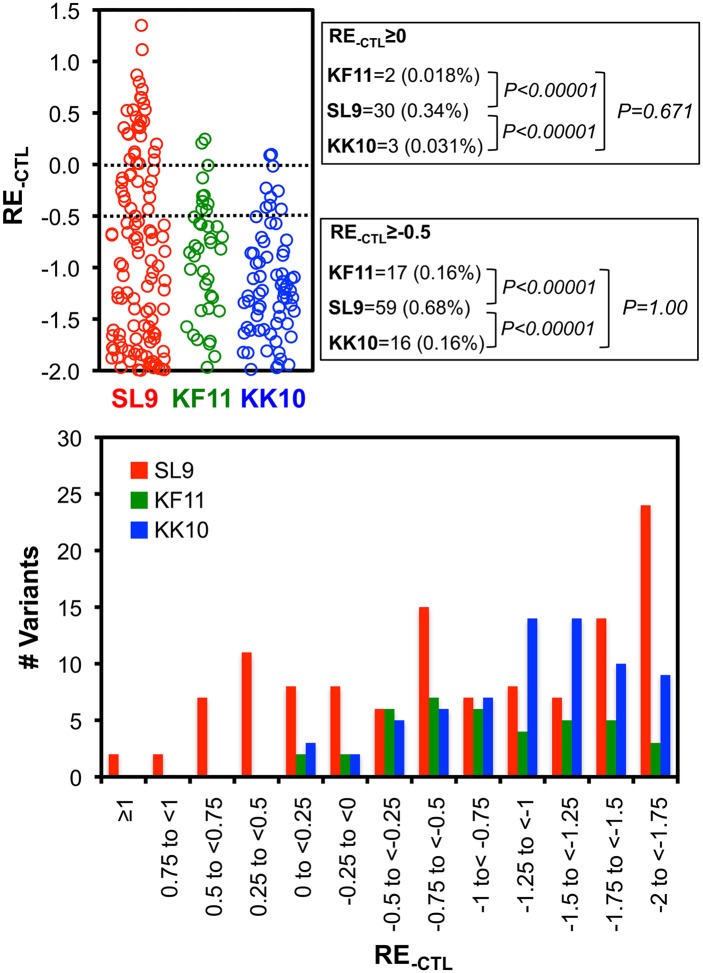
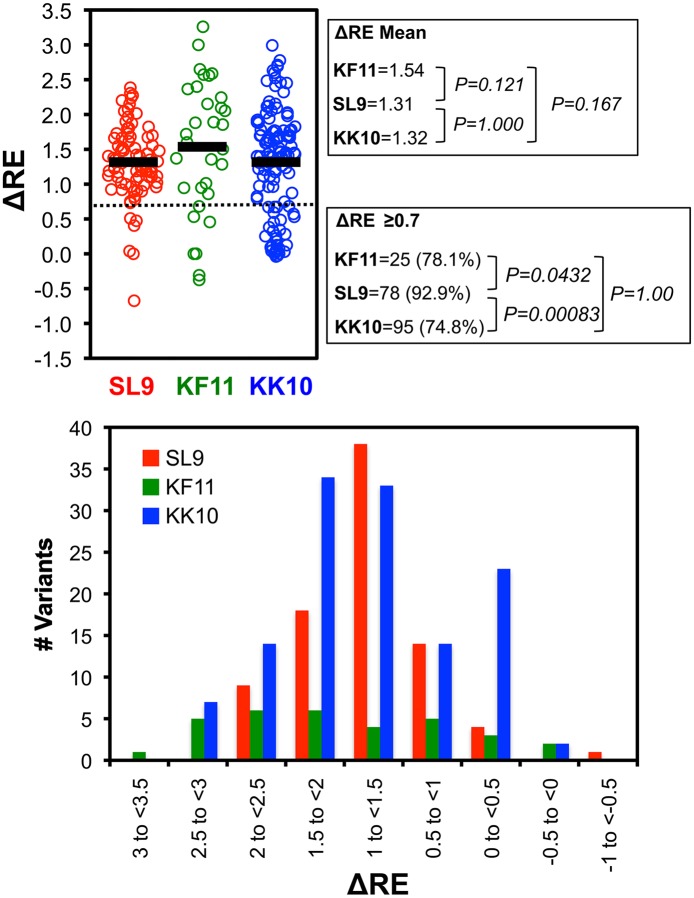
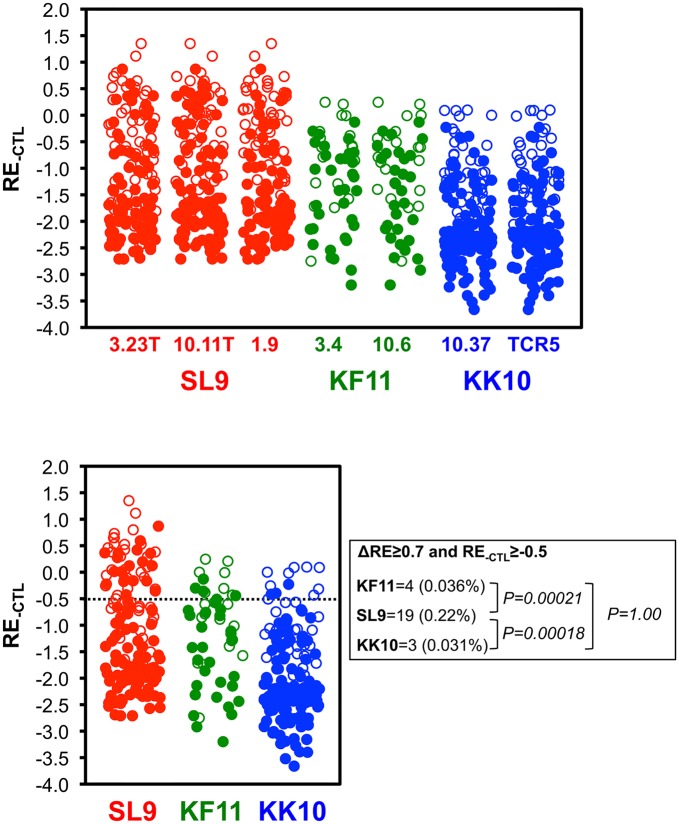
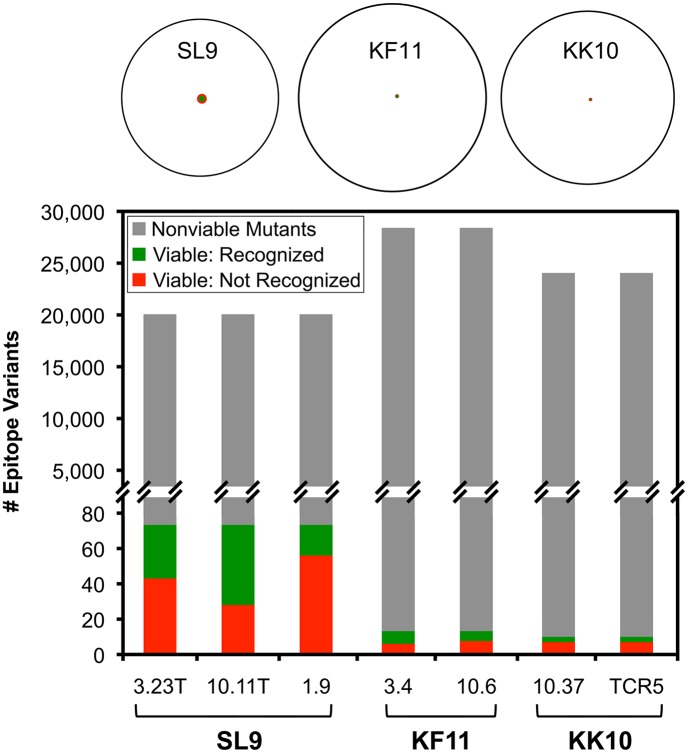
Certain Major Histocompatibility-I (MHC-I) types are associated with superior immune containment of HIV-1 infection by CD8+ cytotoxic T lymphocytes (CTLs), but the mechanisms mediating this containment are difficult to elucidate in vivo. Here we provide controlled assessments of fitness landscapes and CTL-imposed constraints for immunodominant epitopes presented by two protective (B57 and B27) and one non-protective (A02) MHC-I types. Libraries of HIV-1 with saturation mutagenesis of CTL epitopes are propagated with and without CTL selective pressure to define the fitness landscapes for epitope mutation and escape from CTLs via deep sequencing. Immunodominant B57- and B27- present epitopes are highly limited in options for fit mutations, with most viable variants recognizable by CTLs, whereas an immunodominant A02 epitope-presented is highly permissive for mutation, with many options for CTL evasion without loss of viability. Generally, options for evasion overlap considerably between CTL clones despite highly distinct T cell receptors. Finally, patterns of variant recognition suggest population-wide CTL selection for the A*02-presented epitope. Overall, these findings indicate that these protective MHC-I types yield CTL targeting of highly constrained epitopes, and underscore the importance of blocking public escape pathways for CTL-based interventions against HIV-1.
某些主要组织相容性复合体-I(MHC-I)类型与CD8 +细胞毒性T淋巴细胞(CTL)对HIV-1感染的卓越免疫控制相关,但介导这种控制的机制在体内难以阐明。在这里,我们对由两种保护性(B57和B27)和一种非保护性(A02)MHC-I类型呈递的免疫显性表位的适应性景观和CTL施加的限制进行了对照评估。通过对CTL表位进行饱和诱变的HIV-1文库在有和没有CTL选择压力的情况下进行繁殖,以通过深度测序确定表位突变和逃避CTL的适应性景观。免疫显性的B57和B27呈递的表位在合适突变的选择上受到高度限制,大多数可行变体可被CTL识别,而免疫显性的A02呈递的表位对突变高度宽容,有许多逃避CTL的选择且不丧失活力。一般来说,尽管T细胞受体高度不同,但CTL克隆之间的逃避选择有相当大的重叠。最后,变体识别模式表明群体范围内CTL对A*02呈递的表位进行选择。总体而言,这些发现表明这些保护性MHC-I类型产生针对高度受限表位的CTL靶向作用,并强调了阻断公共逃逸途径对于基于CTL的HIV-1干预措施的重要性。