Singh Gunjita, Da Ines Olivier, Gallego Maria Eugenia, White Charles I
Génétique, Reproduction et Dévelopement, UMR CNRS 6293 - INSERM U1103 - Université Cleront Auvergne Campus Universitaire des Cézeaux, Aubiere, France.
PLoS One. 2017 Aug 10;12(8):e0183006. doi: 10.1371/journal.pone.0183006. eCollection 2017.
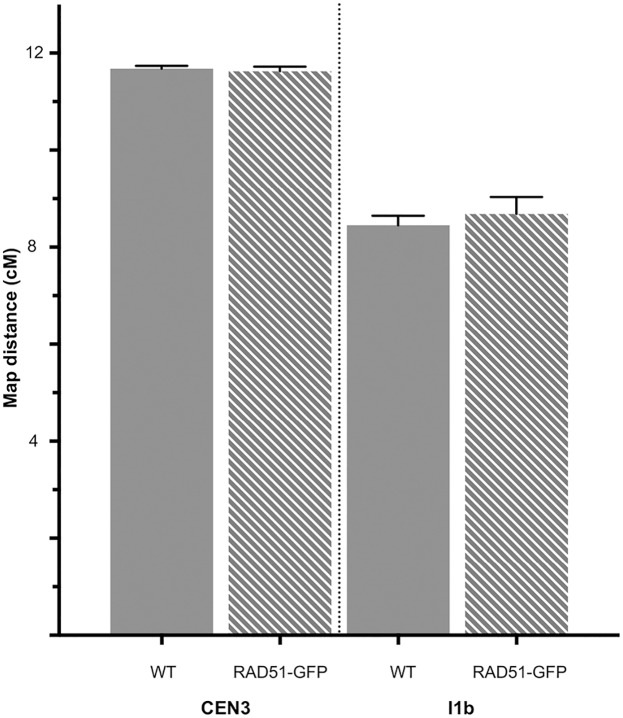
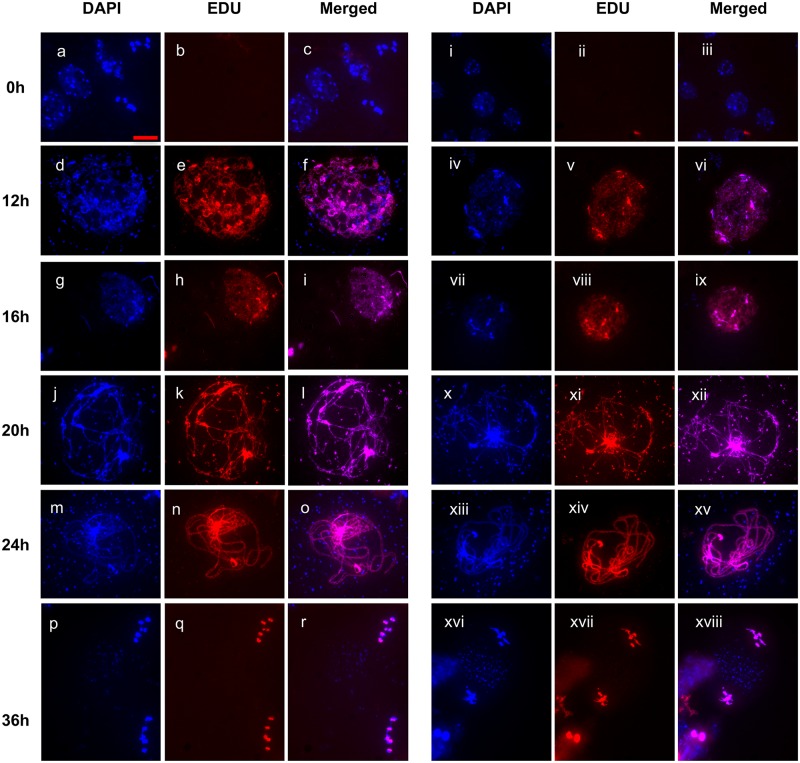
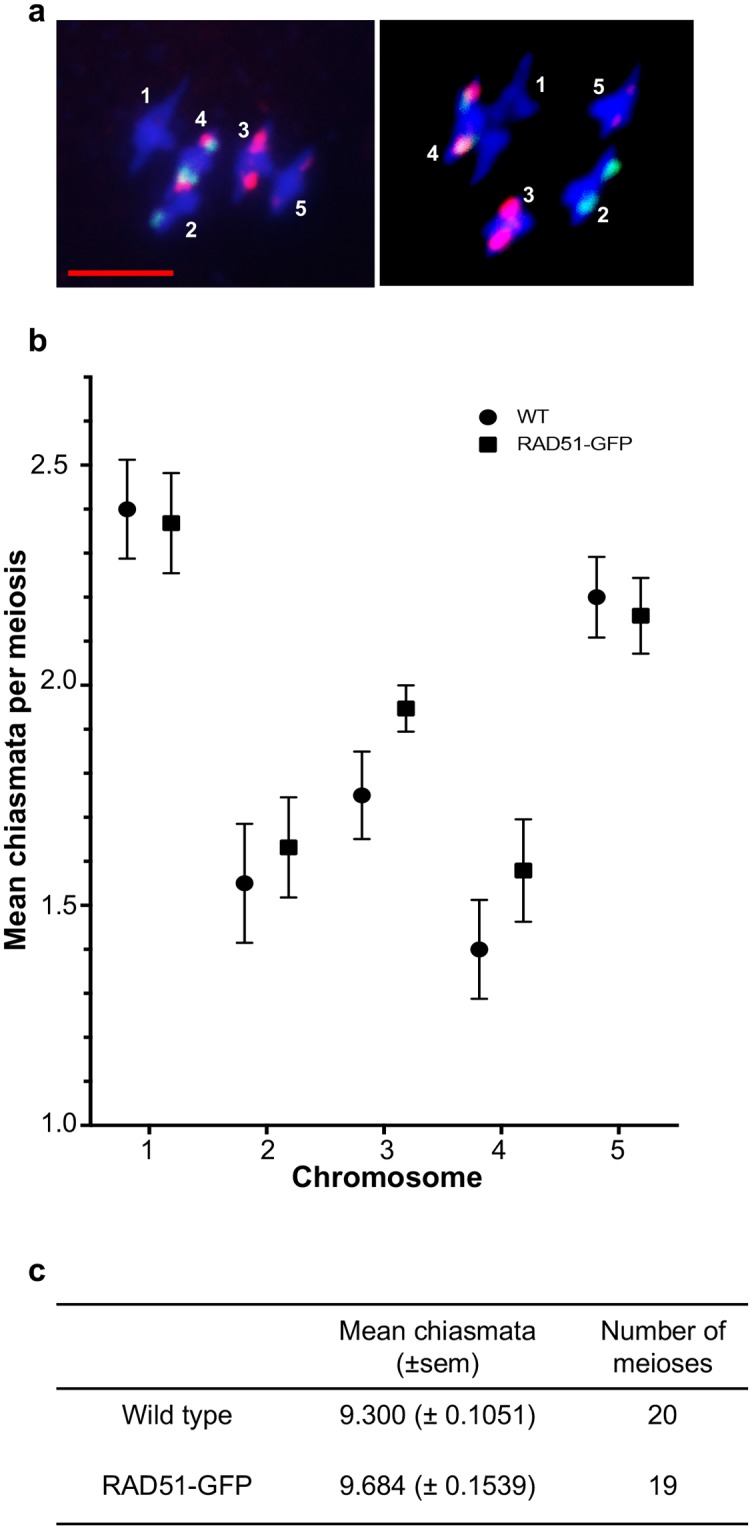
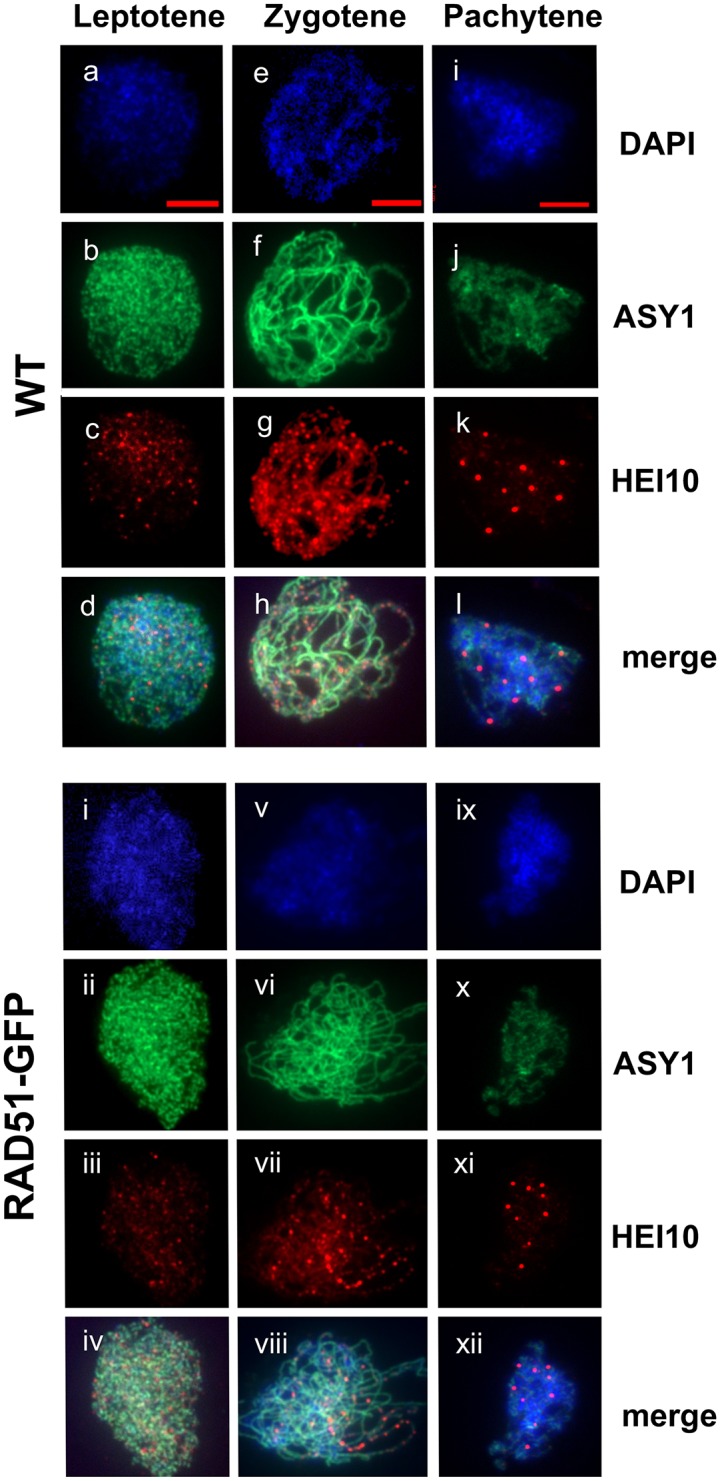
The ploidy of eukaryote gametes must be halved to avoid doubling of numbers of chromosomes with each generation and this is carried out by meiosis, a specialized cell division in which a single chromosomal replication phase is followed by two successive nuclear divisions. With some exceptions, programmed recombination ensures the proper pairing and distribution of homologous pairs of chromosomes in meiosis and recombination defects thus lead to sterility. Two highly related recombinases are required to catalyse the key strand-invasion step of meiotic recombination and it is the meiosis-specific DMC1 which is generally believed to catalyse the essential non-sister chromatid crossing-over, with RAD51 catalysing sister-chromatid and non-cross-over events. Recent work in yeast and plants has however shown that in the absence of RAD51 strand-exchange activity, DMC1 is able to repair all meiotic DNA breaks and surprisingly, that this does not appear to affect numbers of meiotic cross-overs. In this work we confirm and extend this conclusion. Given that more than 95% of meiotic homologous recombination in Arabidopsis does not result in inter-homologue crossovers, Arabidopsis is a particularly sensitive model for testing the relative importance of the two proteins-even minor effects on the non-crossover event population should produce detectable effects on crossing-over. Although the presence of RAD51 protein provides essential support for the action of DMC1, our results show no significant effect of the absence of RAD51 strand-exchange activity on meiotic crossing-over rates or patterns in different chromosomal regions or across the whole genome of Arabidopsis, strongly supporting the argument that DMC1 catalyses repair of all meiotic DNA breaks, not only non-sister cross-overs.
真核生物配子的倍性必须减半,以避免每一代染色体数量翻倍,这一过程通过减数分裂来实现。减数分裂是一种特殊的细胞分裂,其中单一的染色体复制阶段之后是连续两次核分裂。除了一些例外情况,程序性重组确保了减数分裂中同源染色体对的正确配对和分布,因此重组缺陷会导致不育。催化减数分裂重组关键链侵入步骤需要两种高度相关的重组酶,一般认为减数分裂特异性的DMC1催化必需的非姐妹染色单体交叉,而RAD51催化姐妹染色单体和非交叉事件。然而,最近在酵母和植物中的研究表明,在缺乏RAD51链交换活性的情况下,DMC1能够修复所有减数分裂DNA断裂,令人惊讶的是,这似乎并不影响减数分裂交叉的数量。在这项工作中,我们证实并扩展了这一结论。鉴于拟南芥中超过95%的减数分裂同源重组不会导致同源染色体间的交叉,拟南芥是测试这两种蛋白质相对重要性的一个特别敏感的模型——即使对非交叉事件群体有微小影响,也应该会对交叉产生可检测的影响。虽然RAD51蛋白的存在为DMC1的作用提供了必要支持,但我们的结果表明,缺乏RAD51链交换活性对拟南芥不同染色体区域或整个基因组的减数分裂交叉率或模式没有显著影响,这有力地支持了DMC1催化所有减数分裂DNA断裂修复的观点,而不仅仅是非姐妹交叉。